| Головна |
| «« | ЗМІСТ | »» |
|---|
Теоретично допустимо, що кросинговер відбувається як до реплікації хромосом - на стадії двох ниток, так і після - на стадії чотирьох ниток. це підтверджує зошита аналіз, оскільки при цьому підході можливо дослідження всіх чотирьох продуктів кожного мейозу. Одним з найбільш зручних об'єктів для зошитового аналізу є гаплобіонт хлібна цвіль {Neurospora crassa) (Рис. 6.2).
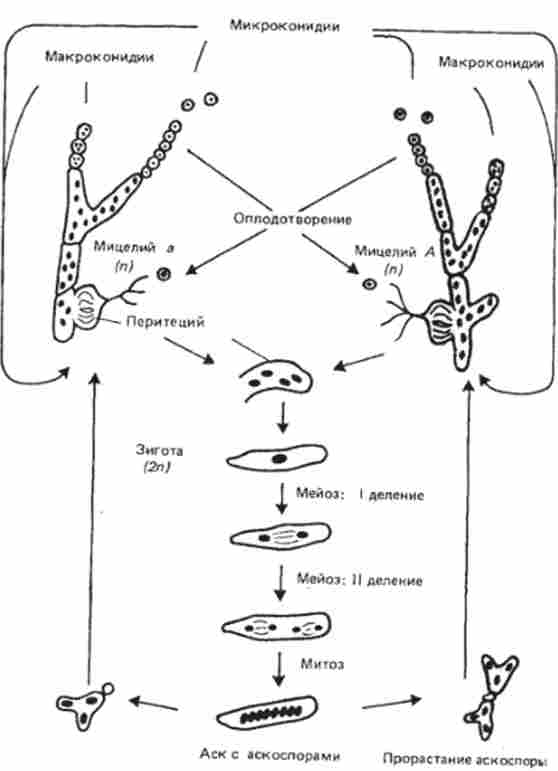
Мал. 6.2. Життєвий цикл хлібної цвілі (Neurospora crassa)
Особливість мейозу у хлібній цвілі в збігу напрямки веретена I і II ділення мейозу з довгою віссю аска, або сумки, в якій розташовуються потім гаплоїдні аскоспори. Чотири гаплоїдних ядра після мейозу ще раз діляться мітотично, в результаті чого в аске в ряд розташовуються чотири пари гаплоїдних спор, а генотип кожної пари суперечка ідентичний.
При тісному зв'язку генів вкрай рідкісний кроссинговер на стадії двох ниток в тетрадах (октадах) нейроспори (рис. 6.3, а, б). Причина їх появи пов'язана з певними типами подвійних обмінів.
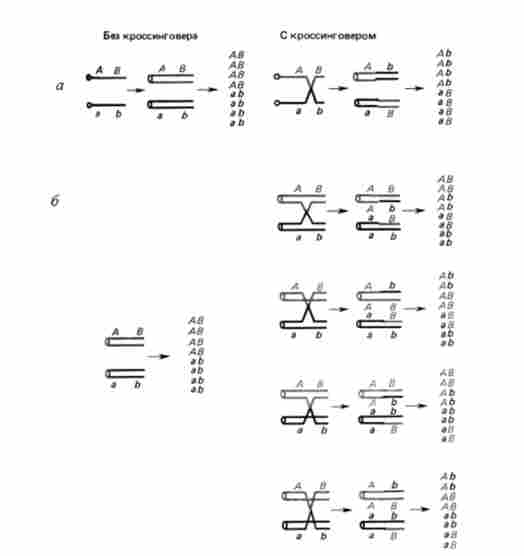
Мал. 6.3. Можливі наслідки кросинговеру у хлібній цвілі {Neurospora crassa) при дигибридном схрещуванні: а - на стадії двох ниток; б - на стадії чотирьох ниток. Гуртки - центромери
Як правило, кроссоверние аскоспори містяться в тетрадах (окта- дах). Наслідки кросинговеру на стадії чотирьох ниток розглянуті на рис. 6.3. Розбіжність центромер у N. crassa орієнтоване по довжині аска, у якого розрізняють базальний і апікальний кінці. Доказ того, що кросинговер йде на стадії чотирьох ниток, можна отримати і при розгляді моногибридного схрещування. При схрещуванні штамів, що розрізняються по аллелям тільки одного гена В / b, завжди є ще один маркер, в якості якого служить напрямок розбіжності центромер при двох розподілах мейозу. Порядок розташування аскоспор, що виникає внаслідок кросинговеру на ділянці ген-центромера, підтверджує висновок про те, що рекомбінація станься на стадії чотирьох хроматид.
У цьому можна переконатися, якщо розглядати тільки розщеплення по гену В / b, дистальному від центромери. У разі кросинговеру на ділянці ген В-центромера не збігаються редукція по центромере і редукція по гену В / b. Редукція по центромере відбувається при мейозі I, а редукція по генетичному факторі - при мейозі II. Редукція по центромере і по гену В / b збігається тільки тоді, коли на ділянці ген-центромера немає кроссін- Говера. Таким чином, завдяки особливостям життєвого циклу нейроспори у неї можна картировать гени по відношенню до Центромера. Для цього потрібно враховувати частоту редукції (розщеплення) з даного гену при мейозі II, про яку можна судити по розташуванню аскоспор в асках.
У тому, що кросинговер відбувається після реплікації хромосом, можна переконатися не тільки на прикладі нейроспори і інших організмів, у яких можливий зошита аналіз. У 1925 р К. Бріджес і І. Андерсон продемонстрували хроматидного кроссинговер у дрозофіли. Для цього вони використовували лінію зі зчепленими А-хромосомами, яка несе також У-хромосому (А х ХУ), аналогічну лінії double yellow. Мухи в дослідах К. Бріджеса і І. Андерсона були гетерозиготних по генам А-хромосоми: f (forked) - роздвоєні, копитні щетинки, g (garnet) - яскраво-червоні очі, v (vermilion) - також яскраво-червоні очі. При схрещуванні самок з таким набором хромосом зі звичайними самцями вони безпосередньо передають дві свої А-хромосоми дочкам, і виживає тільки половина їх потомства. При цьому частина самок у потомстві від такого схрещування виявляються гомозиготними по рецесивним генам А-хромосоми. Очевидно, такі гомозиготи можуть з'являтися тільки в результаті кросинговеру на стадії чотирьох хроматид на ділянці ген-центромера. Дослідивши гомозиготизації по трьом різним генам А-хромосоми, вчені переконалися, що її частота пропорційна їх відстані від центромери, яка знаходиться на самому кінці А-хромосоми (для / - 5,5%, для g - 10%, для v - 16,1%). Цей спосіб аналізу у дрозофіли отримав назву полутетрадного, оскільки кожна враховується в експерименті самка (А х XY) несе дві з чотирьох хроматид бивалента, що утворюється в профазі I мейозу.
Переконавшись, що кросинговер відбувається на стадії чотирьох ниток, звернемося до експерименту Дж. Тейлора (1967), доводить справедливість гіпотези «розрив - возз'єднання». Прознавств ним експеримент - своєрідний варіант зошитового аналізу з використанням цитологічного методу. Німфам коника вводили тимидин, мічений тритієм так, щоб він включався в хромосоми в останньому мітотичного циклу перед мейозом. Завдяки цьому в результаті подальшої премейотічної реплікації кожна хромосома бивалента складалася з міченої і немічених хроматид. Результати реципрокних обмінів між міченими і немічених хроматину були чітко видні на радіоавтографах. Такі обміни представляють лише половину всіх відбуваються, т. К. Обміни тільки між міченими або тільки між немічених хроматину не реєструються.
Частота спостережуваних обмінів відповідала передбаченої виходячи з числа хиазм і відсутності сестринських обмінів. Наприклад, середнє число хіазм, що припадають на найдовшу хромосому, так само 3,67. Оскільки обміни здійснюються на стадії чотирьох хроматид, кожна хіазма дає тільки дві рекомбінантні хроматиди (3,67: 2 = 1,88). Реєструється половина всіх обмінів (1,88: 2 = 0,94). Для цієї хромосоми Дж. Тейлор спостерігав рекомбінантні хроматиди з частотою 0,89.
Зошитовий аналіз незамінний при вивченні множинних обмінів і інтерференції між ними. Для цього розглядають трігібрідное схрещування (ЛВС х abc) по зчепленим генам. З огляду на, що кроссінго- вір відбувається на стадії чотирьох хроматид, можливі три типи подвійних обмінів. це подвійні двуххроматідние обміни, подвійні треххроматід- ні обміни і подвійні четиреххроматідние обміни тільки між НЕ сестринськими хроматидами, наслідки яких генетично помітні (рис. 6.4).
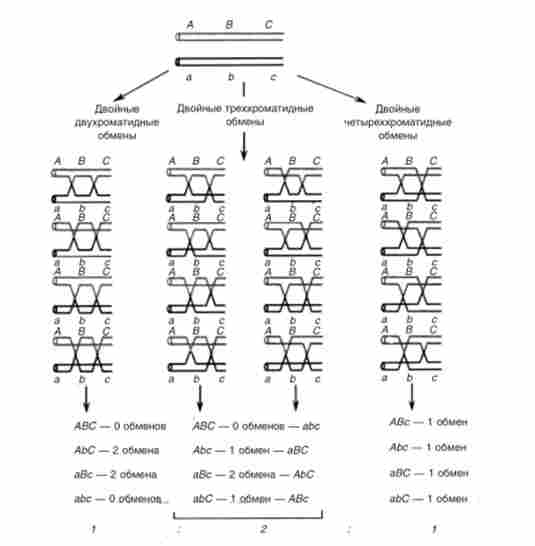
Мал. 6.4. Типи подвійних обмінів, реєстровані при тетрадном аналізі трігнбрідного схрещування у грибів.
Внизу - очікуване співвідношення типів зошити; порядок суперечка не фіксований
Можна ідентифікувати наслідки двох-, трьох- і чотирьох-хроматидного обмінів по присутності в тетрадах аскоспор без обмінів (АВС і abc), З одним обміном (АВС і аЬС або Abc і АВС) і з двома обмінами: (АЬС і АВС).
Розташування аскоспор в даному випадку несуттєво. Якщо припустити, що всі типи обмінів різновірогідні, то слід очікувати появи зазначених трьох типів тетрад в співвідношенні 1: 2: 1.
У 1963 р С. Емерсон статистично обробив представлені багатьма авторами дані по зошитового аналізу подвійного кросинговеру і отримав загальне співвідношення двох-, трьох- і четиреххроматідних обмінів - 0,29: 0,48: 0,23. Це співвідношення близько до 1: 2: 1. Отже, різні типи подвійних обмінів відбуваються практично випадково, т. Е. Хроматиди, які вступили або не набрали один акт рекомбінації, з однаковою ймовірністю беруть участь в повторному акті рекомбінації.
Наведені дані показують, що хроматидного інтерференція Відсутнє. Її відсутність не суперечить існуванню хромосомної, або хіазменной, інтерференції. Це означає, що один обмін ускладнює другий і наступні обміни на сусідніх ділянках, але якщо подвійний кросинговер все ж відбувається, то всі чотири хроматиди залучаються до нього равновероятно.
Мабуть, важливу, але до кінця не з'ясованою роль в мейотіче- ському кроссинговере грає сінаптонемний комплекс (СК). У різних організмів здатність до кросинговеру корелює з утворенням СК. Наприклад, він формується у самок D. melanogaster з нормальним кроссін- Говера, але не у самців, у яких кроссинговер пригнічений. При утворенні СК невеликі ділянки ДНК кон'югується хромосом залучаються всередину центральної зони СК. Передбачається, що саме на цих ділянках ініціюються межхроматідние обміни. У рослин відомі ахіазматі- етичні мутанти, у яких зміна одного гена призводить до втрати СК, відсутності хиазм і придушення кросинговеру.
Нами розглянуті обміни тільки між НЕ сестринськими хроматину. Для з'ясування питання про можливість і частоті обмінів між сестринськими хроматину в мейозі потрібні спеціальні генетичні підходи.
Таким чином, кроссинговер відбувається на стадії чотирьох ниток - в профазі мейозу. В результаті цього утворюються хіазми, які найзручніше спостерігати на стадіях діплотени і діакінеза.