| Головна |
| «« | ЗМІСТ | »» |
|---|
Нирки - це основний орган, що забезпечує сталість обсягу і складу внутрішнього середовища, і перш за все крові. Реалізацію гомеостатичної функції здійснюють спеціальні рефлекторні системи, що включають спеціалізовані рецептори (оцінюють об'єм циркулюючої крові - волюмо- рецептори, осморецептори, іонорецептори), системи переробки інформації (нервові центри) і «командні» структури, що опосередковують свій вплив на тканину нирки за рахунок еферентних нервів і гормональних впливів.
Осморегуляція. При надмірному вмісті води в крові концентрація осмотично активних речовин знижується і осмотичний тиск крові падає, що вловлюється як центрально розташованими осморецепторами в області супраопті- чеських ядер гіпоталамуса, так і периферичними рецепторами нирок, печінки, селезінки. В результаті істотно знижується вироблення антидіуретичного гормону (АДГ) і його надходження в кров з нейрогипофиза, що призводить до посилення виділення води ниркою за рахунок утворення менш концентрованої сечі. Зневоднення організму призводить до збільшення концентрації осмотично активних речовин. В результаті активуються осморецептори, посилюється продукція і виведення АДГ, зростає канальцевая реабсорбція води, включається механізм концентрації сечі, зменшується сечовиділення.
Волюморегуляція. Обсяг внутрішньосудинної рідини оцінюється рецепторами, що реагують на зміну тонусу судинної стінки. В області низького тиску від рецепторів лівого і правого передсердя імпульси передаються в ЦНС по аферентні волокнах блукаючого нерва. При збільшенні об'єму крові (внутрішньосудинної циркулюючої рідини) підвищується імпульсація з волюморецепторов, що викликає екскрецію натрію і води. Одночасна активація волюмо- і осморецепторов призводить до посилення виведення води, зменшуючи її реабсорбцію.
Іонорегуляціі. Дані про наявність рецепторів, які оцінюють вміст натрію (введення в область III шлуночка гіпертонічного розчину хлориду натрію викликає антідіуреза, а гіпертонічних розчинів цукру такого ефекту не викликає), вказують на можливість виборчої оцінки концентрацій окремих іонів і адекватної реакції видільної системи в ході рефлекторних реакцій. Відомі гормони, що регулюють реабсорбцію і секрецію окремих іонів в різних відділах нефрона. Реабсорбція натрію зростає в дистальних відділах ниркових канальців під впливом гормону надниркової залози аль- достерона, який відносно калію виявляється речовиною, що стимулює його видалення з сечею. Відомо, що альдостерон зв'язується з клітинами дистального і збірного сегментів нирок і може збільшувати реабсорбцію натрію шляхом синтезу РНК і одного або більше специфічних білків. В результаті може безпосередньо стимулюватися механізм активного транспорту натрію, збільшуватися проникність для натрію мембрани клітин, звернених в просвіт канальця, або стимулюватися клітинний метаболізм і зростати поставка енергії для транспортних процесів. Виділення калію зменшується під дією інсуліну.
Зниження концентрації кальцію в крові викликає виділення паращитовидних залозами паратгормона, завдяки якому збільшується реабсорбції цього іона в ниркових канальцях. Дистальний відділ нефрона є головний ділянку, де здійснюється дію ПТГ. Гіперкальціе- мія стимулює секрецію щитовидною залозою тірокальціто- нина, який знижує концентрацію кальцію в крові за рахунок посилення його екскреції ниркою і мобілізації в кістковій тканині. У ниркових канальцях регулюється також рівень реабсорбції магнію, хлору, фосфатів, сульфатів та інших іонів.
Регуляція кислотно-лужної рівноваги. Концентрація іонів водню в сечі може істотно змінюватися в бік зниження (до 4) або зростання (до 8,0), що покликане забезпечити сталість pH плазми крові на рівні 7,36. Механізм видалення іонів водню (закислення сечі) забезпечується процесами секреції цього іона в просвіт канальців. Наявність в апікальній плазматичній мембрані ферменту карбоангідрази забезпечує гідратацію двоокису вуглецю і утворення вугільної кислоти, яка відразу дисоціює на іон водню і двокарбонатний іон.
Водневі іони секретуються в канальцеву рідину, а утворені бікарбонатні іони повертаються в плазму, т. Е. Секреція Н+ відповідає освіті бікарбонатів в плазмі. Вступник з плазми крові в ході ультрафільтрації ИаНС03 взаємодіє з іонами водню, секретуватися клітинами, що призводить до утворення діоксиду вуглецю (рис. 8.10). В просвіті канальців іони водню здатні зв'язуватися і з фосфатами, і з іншими аніонами, що призводить до екскреції з сечею тітруемих кислот, а в плазмі крові збільшується рівень підстав. Крім того, аміак, що виділяється в кров як кінцевий продукт білкового обміну, також здатний зв'язуватися з іонами водню з утворенням іона амонію. Аміак пов'язує кислі продукти, що виділяються з сечею, утворює амонійні солі і, віддаляючись за межі організму, заміщає натрій і калій. Аміак, який використовується для нейтралізації кислої сечі, утворюється в нирках з амінокислоти глутаміну за рахунок функціонування ферменту глутаміна- зи, яка активується в разі необхідності корекції реакції сечі. В результаті нирки стабілізують концентрацію іонів водню в плазмі крові: при зниженні резервної лужності крові виділяється кисла сеча, а при алкалозі - більш лужна.
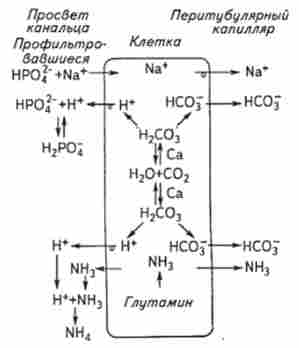
Мал. 8.10. Освіта титруемой кислотності і іонів амонію в нирковому канальці
Таким чином, кількість екскретуватися водневих іонів з сечею - сумарна екскреція Н+, може бути представлена наступним виразом:
Сумарна екскреція Н+ = 14Н4 + титруемая кислотність НС03 сечі.
Зазвичай загальна кількість екскретуватися іонів водню дорівнює водневої навантаженні. Сумарна екскреція Н+ може бути відсутнім або мати негативне значення, що відбувається при бикарбонатной навантаженні (високому вмісті цитратов в кормі), цитрати метаболізуються до бікарбонатів і сеча набуває лужну реакцію.
Екскреція. Через нирку виводиться велика частина кінцевих продуктів обміну речовин. Особливо це важливо для метаболізму білків і нуклеїнових кислот, в ході якого утворюються різні продукти азотистого обміну. В процесі перетворення амінокислот аминогруппа (-N112) видаляється шляхом дезамінування і утворює аміак (14Н3), Який виділяється у вигляді амонійних солей або використовується в синтезі азотовмісних сполук - сечовини і сечової кислоти.
Синтез сечовини здійснюється в послідовних ферментативних реакціях, що складають орнітіновий цикл. Аміак і двоокис вуглецю, конденсуючись з фосфатом, утворюють карбо- моілфосфат, який використовується для синтезу цітруміна з орнитина. Приєднання ще однієї молекули аміаку з аспарагінової кислоти призводить до утворення амінокислоти аргініну. У присутності аргінази аргінін розпадається на сечовину і орнітин; з орнитина синтезується нова молекула цитруллина, і весь цикл повторюється. Виділення сечової кислоти переважає у тварин, які змушені економити воду (рептилії і птахи). Оскільки сечова кислота і її солі дуже погано розчиняються у воді і реабсорбція води призводить до випадання щільного осаду, то для виведення азотистих екськретов витрачається дуже мало води, т. Е. Для виведення кінцевих продуктів обміну її потрібно значно менше, і це особливо важливо для зниження маси тіла птахів, що важливо для польоту.
Нуклеїнові кислоти містять дві групи азотистих сполук: пурини (Аденін і гуанін) і піримідинів (Цитозин і тимін). У деяких тварин пурини виділяються у вигляді сечової кислоти, у інших - пуринова структура розщеплюється до проміжних з'єднань або аміаку. Людина і вищі мавпи через відсутність ферменту уриказа не здатні метаболизировать сечову кислоту і виділяють її з сечею, решта ж ссавці володіють цим ферментом і перетворять сечову кислоту в аллан- тоін. Серед собак у долматінского дога виявлено інтенсивне виділення сечової кислоти, але не через відсутність уриказа, а як наслідок спадкового ниркового дефекту, що перешкоджає канальцевої реабсорбції, а, можливо, екскреції сечової кислоти шляхом активного канальцевого транспорту.
Гормонопоез. Нирки виробляють ряд фізіологічно активних речовин, що надходять в кровотік і беруть участь в системних реакціях організму, що підтверджує їх ендокринну функцію. Гранулярні клітини юкстагломерулярних нефронів здатні синтезувати і виділяти в кров ренін - протеолітичний фермент, який в плазмі крові отщепляет від ангіотензиногена декапептид ангіотензин I, потім від ангіотензину I відщеплюються ще дві амінокислоти, і утворюється досить активне з'єднання ангіотензин II. Ангіотензин II викликає звуження судин, стимулює секрецію альдостерону і антидіуретичного гормону і підсилює ниркову реабсорбцію натрію, а також бере участь у формуванні почуття спраги (рис. 8.11).
Секреція альдостерону покликана підвищувати реабсорбцію натрію і втрату калію в дистальному відділі нефрона. За принципом зворотного негативного зв'язку секреція реніну відзначено зниження ангиотензином II і антидиуретичним гормоном, але разом з цим секреція реніну істотно залежить від стану барорецепторів нирки, вмісту натрію в початковій частині канальців і симпатичних нервових впливів. Таким чином, рівень активності реніну плазми знижується при збільшенні обсягу позаклітинної рідини, сольовий навантаженні, надлишку мінералкор- тікоідов і гіперкаліємії. Підвищення рівня змісту рені-
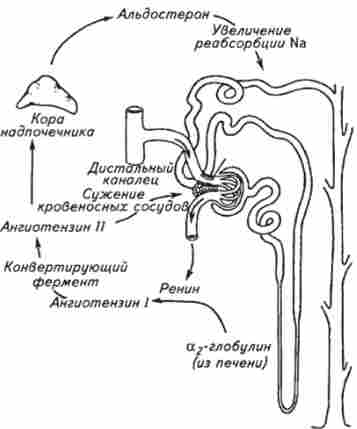
Мал. 8.11. Схема взаємодії системи ангіотензину та альдостерону
на розвивається при зменшенні об'єму позаклітинної рідини, обмеження солі, втрати рідини (блювання, пронос, діуретики), гіпокаліємії, надлишку катехоламінів і при збільшенні вмісту в плазмі крові ренін-субстрату за рахунок посиленого синтезу в печінці. Глюкокортикоїди і статеві стероїдні гормони здатні підвищити рівень ангіотензиногена до концентрацій, що активують ренін.
Каллікреіновой система багато в чому схожа з реніновою. Каллі- креін є пептідази, що впливає на 012-глобулін (кініген) з утворенням кініну. Термін 1 кинин відноситься до трьох біологічно активним пептидів: метіоніл-гліцил-брадикинину, лізил-брадикинину і брадикинину. Кініни - це потужні вазодилататори. Ниркова каллікреіновой система є локальною гормональною системою, залученої в регуляцію кровотоку і екскрецію натрію. Калікреїн синтезується в кірковій речовині нирки і забезпечує продукцію потужного судинорозширювальної речовини каллидин. Відомо, що калликреин нирок звільняється і надходить у кров під впливом високого артеріального тиску, ацетилхоліну, простагландину, дофаміну, низьких доз норадреналіну, ангіотензину II, минералкортикоидов і при швидкому збільшенні об'єму рідини, що циркулює в організмі.
У мозковій речовині нирки утворюються простагландини, в тому числі і простагландин А2, який називають також медулліном, а також простациклін, або Р§12. Простагландини здатні впливати на периферичний судинний опір за рахунок їх дії як вазодилататорів, натрийуретичних речовин, інгібіторів адренергічних медіаторів і зміни чутливості клітин до антидіуретичного гормону. Нирковий синтез Р§12 стимулюється ангітензіном II, брадикініном і вазопресином. Збільшення синтезу відбувається за рахунок підвищеного звільнення арахідонової кислоти з ниркових ліпідів під впливом ферменту ацілгідролази.
Регуляція кількості еритроцитів здійснюється одним з гормонів нирки - еритропоетином, який представляє собою глікопротеїн з м. М. 40000. Стимулом для підвищеного синтезу еритропоетину тканинами нирки є зменшення напруги в ній кисню.
Клітини нирки беруть участь в метаболізмі вітаміну Е: що утворюється в печінці прогормон вітамін 03 нирки витягають з кровотоку і перетворять в ряд диокси-похідних (найбільш важливими є 1,25-диокси-Оз і 24,25-диокси-02). 1,25-диокси-03 як вельми активний стероїдний кальціемічний гормон збільшує транспорт кальцію і фосфору в кишечнику, регулює реабсорбцію кальцію в ниркових канальцях і підвищує мінеральну резорбцію кісток.
Метаболізм. Крім забезпечення власних енергетичних витрат при виконанні специфічних функцій нирки беруть участь в обміні речовин, покликаному підтримувати гомеостаз. У нирці є система новоутворення глюкози - глюконеогенез. При тривалому голодуванні з нирок в кровотік може надходити до половини загальної кількості глюкози. Вступники при фільтрації в просвіт канальців низькомолекулярні білки і пептиди розщеплюються клітинами канальців до амінокислот, які через базолатеральную мембрану повертаються в кровотік. У тканині нирки синтезується один з важливих компонентів біомембран фосфатіділінозіт, а вільні жирні кислоти включаються до складу тригліцеридів і фосфоліпідів, які кровотоком поставляються до місць депонування або утилізації.