| Головна |
| «« | ЗМІСТ |
|---|
Вперше припущення про рекомбінаційному механізмі хромосомнихаберацій висловив А. С. Серебровський (1929). Він виходив з реципрокного характеру транслокаций, рекомбінаційного походження дуплікацій і делеций, а також передбачав необхідність реципрокного обміну в разі інверсій.
Було відмічено також, що точки розривів при транслокації у дрозофіли переважно локалізуються в районах ектопічеського спарювання, т. е. в районах хромосом, час від часу сінапті- рующих на препаратах слинних залоз. Це вказувало на існування в негомологічних хромосомах ділянок часткової гомології і дозволяло шукати рекомбінаційний механізм хромосомних перебудов. Безсумнівний успіх в цьому напрямі був досягнутий тільки з відкриттям мігруючих елементів геному і в зв'язку з застосуванням техніки рекомбінантних ДНК для їх виділення і вивчення.
Одним з найбільш важливих моментів, пов'язаних з вивченням мігруючих елементів, є виявлення частих хромосомних перебудов поблизу точок впровадження / 5-елементів та транспозони. Це справедливо як для бактерій, так і для одноклітинних і багатоклітинних еукаріот. В даний час з урахуванням переміщення мігруючих елементів і механізму звичайної (гомологичной) рекомбінації, а також рекомбінації між / 5-елементами вдається пояснити всі типи відомих хромосомних перебудов.
Делеції і дуплікації можуть відбуватися, якщо два мігруючих елемента в одній і тій же хромосомі однаково орієнтовані. Тоді рекомбінація по гомології між цими елементами після реплікації між сестринськими хроматидами або гомологічними хромосомами призведе до дуплікації (тріплікаціі) і делеції в якості реііпрокних продуктів рекомбінації (рис. 9.12, а). Делеції можуть виникати і в результаті рекомбінації двох гомологічних елементів, розташованих в одній хромосомі і однаково орієнтованих (рис. 9.12, б). При цьому Дуплікація відсутня.
Інверсії виникають в разі протилежної орієнтації двох гомологічних елементів в одній хромосомі (рис. 9.12, в).
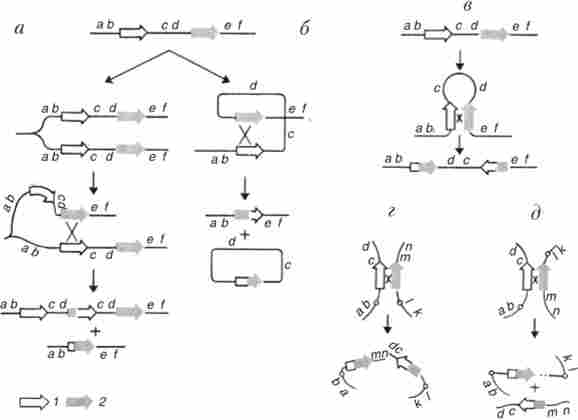
Мал. 9.12. Механізм виникнення хромосомних перебудов, опосередкованих мігруючими елементами: а - делеции і дуплікації; б - делеции без дуплікацій; в - інверсії; г - транслокации; д - освіту мостів і фрагментів в анафазе 1:
1 - разі основний мігруючого елемента, 2 - копія мігруючого елемента, яка перемістилася в нове місце
Транслокації можуть бути результатом рекомбінації двох копій одного і того ж елемента, інтегрованих в різні хромосоми і розташованих однаково в напрямку від центромери або до центромере (рис. 9.12, г).
Мости і фрагменти, часто спостерігаються в анафазе при індукції хромосомних аберацій іонізуючим випромінюванням, - результат рекомбінації копій одного і того ж мігруючого елемента в негомологічних хромосомах, по-різному орієнтованих до центромере: в одній хромосомі від центромери, в іншій - до центромере (рис. 9.12, д).
Більшість перерахованих тут рекомбінаційних механізмів виникнення хромосомних аберацій продемонстровані в експериментальній роботі з бактеріями і дріжджами. Мігруючі елементи здатні захоплювати і переносити на нове місце гени, поруч з якими вони розташовуються. За образним висловом Р. Б. Хесин, «потрапивши в погану компанію, гени з" добропорядних "перетворюються в" бродяг "» (мінливість геному). Тим самим здійснюється Дуплікація окремих генів, необхідна для дивергенції генетичного матеріалу, т. Е. Виникнення генів з новими функціями. Крім того, повтори однакових або схожих ділянок генетичного матеріалу самі по собі створюють умови для рекомбінації по гомології між генами, розташованими в негомологічних ділянках генетичного матеріалу. Подібна рекомбінація відбувається значно рідше, ніж повністю гомологічних рекомбінація - кросинговер, але вона також пов'язана з ініціації рекомбінацію конверсією. Це характерно для дріжджів-сахароміцетів, що мають два однакових гена his 3: один на своєму місці в хромосомі XY, а інший - внесений з плазмидой в результаті інтегративної трансформації. другий ген his 3 інтегрують в іншу частину генома завдяки рекомбінації плазміди з ту - елементом, який вона також несе. За допомогою такої моделі демонструється конверсія між негомологічної хромосоми. Аналогічний результат був отриманий і для різних генів дріжджів з високим рівнем гомології нуклеотидних послідовностей: сус 1 на сус 7, що кодують з-1 і з-2-цитохроми С. У іншого виду дріжджів «негомологічних» конверсія показана між генами, що кодують дуже близькі за структурою тРНК. У рідкісних випадках «негомологічних» конверсія супроводжується реципрокного транслокації.
Розглянуті способи виникнення повторів в геномі за участю мігруючих елементів слід назвати первинними дуплікації, т. е. дуплікації, що виникають de novo. Очевидно, існує велика група так званих вторинних дуплікацій, прикладами яких можуть бути повтори, які виникають внаслідок кросинговеру в гетерозиготних інверсіях і транслокаціях, а також внаслідок нерозходження хромосом і полиплоидии.
Події, що змінюють структуру хромосом в геномі, традиційно пов'язували з еволюційними перетвореннями генетичного матеріалу. Дуплікації поставляють матеріал для творення нових генів в процесі природного відбору. Інверсії і транслокації сприяють генетичної ізоляції нових форм в процесі їх внутрішньовидової дивергенції. У той же час механізм виникнення хромосомних перебудов довгий час не був відомий, і аберації хромосом вважалися нерегулярними подіями, про що говорить сам термін «аберація», т. Е. Відхилення від норми. Розкриття механізму рекомбінації як основного джерела перебудов хромосом змушує по-новому побачити проблему співвідношення стабільності і лабільності генетичного матеріалу.
Хромосомні перебудови, традиційно розглядаються як один з типів мутації, виявилися пов'язаними з процесами рекомбінації.
Подальше вивчення механізму перебудов хромосом дозволить перетворити цей розділ генетики в область спрямованої перебудови геномів, яка по праву отримає найменування хромосомної інженерії.