| Головна |
| «« | ЗМІСТ | »» |
|---|
кожен таламус (Див. Рис. 8.1; 8.2) являє собою яйцевидне освіту довжиною приблизно 4 см. З латеральної боку таламус межує з хвостатим ядром (див. Параграф 9.2), відділяючись від нього кінцевої смужкою {Stria terminalis). Медіальні поверхні таламуса утворюють бічні стінки верхньої частини III шлуночка. Між цими стінками знаходиться межбугорное зрощення (сіра речовина), що з'єднує правий та лівий таламуса. Передній кінець таламуса кілька загострений, а задній розширений і потовщений.
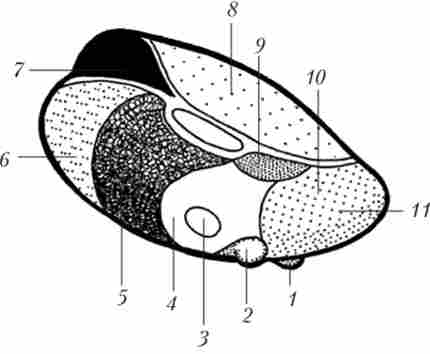
Мал. 8.2. Схема розташування ядер лівого таламуса:
Основна маса таламуса представлена сірим речовиною, згрупованих в ядра (приблизно від 40 до 150 за різними класифікаціями). Більшість ядер таламуса прийнято називати абревіатурами з латинських букв, складених з латинської назви ядра.
В ядрах таламуса відбувається не тільки перемикання інформації, але і її обробка. Одна з основних особливостей цієї обробки полягає у виборчому проведенні інформації в кору великих півкуль. Іншими словами, таламус виконує роль фільтра, пропускаючи в кінцевий мозок або дуже значимі (сильні, нові) сигнали, які сигнали, пов'язані з поточною діяльністю кори великих півкуль. Таким чином, таламус є однією з ключових структур, що забезпечують і підтримують процеси уваги. Для багатьох ядер таламуса, особливо проекційних, характерна присутність гломерул, що говорить про складні процеси аналізу інформації.
Основні класифікації таламичних ядер пов'язані або з їх розташуванням, або з їх функцією. Сіра маса таламуса розділяється медуллярном пластинками (Прошарками білої речовини) на кілька ядерно них груп - передню, медіальну, латеральну, задню і ядра середньої лінії (область межбугорного зрощення і околожелудочкового відділи).
Класифікація ядер таламуса. Більш детально ми розглянемо класифікацію ядер таламуса, засновану на їх функціях та організації зв'язків. З цієї точки зору все таламичні ядра ділять на проекційні, асоціативні і неспецифічні.
Проекційні ядра - це переключательние (релейні) ядра, які отримують входи від дуже обмеженої кількості внеталамічних структур. Волокна з цих структур утворюють синапси на нейронах проекційних ядер, а аксони останніх проводять імпульсацію в певні локальні області кори великих півкуль, що відповідають за певні функції. У свою чергу кожне проекційне ядро отримує спадні волокна з власної проекційної корковою зони. Проекційні ядра поділяються на сенсорні, рухові і лимбичні.
сенсорні (чутливі) ядра забезпечують швидке проведення сенсорної аферентації від конкретних сенсорних систем в первинні проекційні зони кори великих півкуль. Шляхи від всіх рецепторів (за винятком нюхових) проходять через таламус і мають там свої представництва. Головними сенсорними ядрами таламуса є:
Рухові (моторні) проекційні ядра таламуса теж розташовані в його нижній бічній частині перед вентробазального комплексом, тому їх часто називають вентролатерального ядрами. Це два великих ядра VL (П. Ventralis lateralis) і VA (П. Ventralis anterior). Афферентамі цих ядер є структури, пов'язані з організацією рухів, такі як зубчасте ядро мозочка, блідий шар (ядро кінцевого мозку), вестибулярні ядра, чорна субстанція. Ефферентов йдуть в моторну (поле 4) і премоторную (поле 6) кору.
Для ядер вентробазального комплексу і рухових ядер характерна сомаготонія (топографічне представництво поверхні тіла або м'язів).
лимбичні ядра часто називають передніми ядрами таламуса через їх розташування. Це ядра AV (П. Anteroventralis), AD (п. anterodorsalis) Я є (П anteromedialis). Вони входять в ЛС мозку (див. Параграф 9.4) і проводять сенсорну інформацію в лимбичні відділи кори великих півкуль, головним чином в поясний звивину (рис. 9.5). Основні афференти до цих ядер приходять від маміллярних тел гіпоталамуса, сюди приходить також частина волокон зводу (див. Параграф 8.2).
на асоціативних ядрах таламуса закінчуються волокна не від однієї, а відразу від декількох сенсорних систем, а також від інших ядер таламуса і кори великих півкуль. Це забезпечує їх участь в інтеграційних функціях головного мозку, т. Е. В об'єднанні різних видів чутливості. Ці ядра посилають свої волокна в асоціативні зони кори великих півкуль (див. Параграф 9.3). Дорсальні ядра - еволюційно молоді відділи проміжного мозку. Їх формування йде паралельно розвитку вищих асоціативних центрів кори.
Асоціативні ядра LP (П. Lateralis posterior), LD (П. Lateralis dorsalis) і Pul (pulvinar, подушка) разом з зонами їх проекцій в тім'яній ділянці кори великих півкуль розглядають як таламо-париетальную асоціативну систему (Lobus parietalis, тім'яна частка кори), функції якої пов'язані з промовою, а також з розпізнаванням образів і схеми тіла. Окремо треба відзначити, що асоціативне ядро подушка також тісно пов'язане із зоровою системою. Воно отримує афференти від ЛКТ, верхніх горбків четверохолмія, зорової кори, а саме посилає волокна в зорову кору (поля 17, 18, 19). Тому подушку називають іноді зоровим асоціативним ядром.
Медіодорсальнос, або дорсомедіальних, ядро MD {П. medialis dorsalis) Має дуже багато аферентів. Воно отримує волокна від сенсорних і неспецифічних ядер таламуса, від ядра кінцевого мозку мигдалини, від гіпоталамуса, від гіпокампу, від орбітальної і скроневої кори і ін. Характерним для цього ядра є те, що його коркові ефферентов йдуть в лобові асоціативні зони кори, що призвело до формування уявлень про таламо-фроітальіой асоціативної системі. Функції цієї системи остаточно неясні, але в цілому їх можна визначити як формування складних поведінкових актів і контроль емоційних станів.
неспецифічні (медіальні) ядра таламуса зазвичай розглядаються як ядра РФ, що здійснюють зв'язок з ретикулярними ядрами стовбура. Вони отримують афференти від великого числа утворень і посилають дифузні проекції па великі області кори, впливаючи на рівень її активації.
До неспецифічним відносяться ядра середньої лінії, розташовані в його внутрішньої частини, наприклад ядро СМ (п. centralis medialis), інтраламінарние ядра, що лежать серед волокон медуллярной пластинки (lamina, пластинка). До останніх належать, наприклад, велике ядро - серединний центр, або сем (icentrum medianum), І що лежить медіально від нього парафасцікулярное ядро PaF (П. Parafascicularis). Сем і PaF беруть участь в передачі повільних дифузних компонентів больових відчуттів (див. Гл. 15).
Неспецифічним є і ретикулярное таламичне ядро Ret ((Rt), п. reticularis thalami), Що відноситься до латеральної групи ядер. Це гальмівне ядро, що обмежує активацію інших таламичних ядер.