| Головна |
| «« | ЗМІСТ | »» |
|---|
Завдяки цілеспрямованій селекції значно підвищився генетичний потенціал тварин за багатьма господарсько корисним ознаками. Разом з тим все частіше виникають проблеми, пов'язані з плодовитістю тварин і резистентністю їх до хвороб. Досвід показує, що ці проблеми неможливо вирішити тільки за рахунок поліпшення годівлі, технології утримання або засобами ветеринарної терапії. На практиці лікарські засоби і препарати, що підвищують імунітет, дуже часто виявляються недостатніми для лікування і профілактики хвороб.
Очевидно, важливе значення спадковості в прояві резистентності або сприйнятливості тварин до певних хвороб, стійкості до несприятливих факторів зовнішнього середовища. Велике значення має генетична профілактика хвороб всередині виду, в якому знаходяться індивідууми, здатні жити зі збудниками хвороби, в той час як інші хворіють. В результаті накопичення сприятливо діють генів резистентних індивідуумів можна створити резистентні до хвороб лінії і породи.
В даний час використовуються генетичні методи для пошуку і аналізу причин, що обумовлюють зниження рівня репродуктивної функції та життєздатності, поширення аномалій, а також ведеться розробка науково обґрунтованої системи їх профілактики.
Встановлено, що багато форм патології тварин мають генетичну основу і пов'язані з мутаціями і рекомбінаціями спадкового матеріалу - генів і хромосом.
Роль мутацій і рекомбінації генів у виникненні патології у тварин. Мутації, що представляють собою стійкі зміни в структурі ДНК, хромосом і кількісний склад кариотипа, постійно і з певною частотою виникають в популяціях тварин.
Фенотипічно мутації нерідко виявляються в формах вроджених вад (аномалій), в смертності, зниження життєздатності і стійкості до хвороб, порушення репродуктивної функції. У популяціях сільськогосподарських тварин в процесі тривалого їх існування накопичено певний вантаж шкідливих рецесивних мутацій і аберацій хромосом. Для профілактики поширення шкідливих мутацій необхідна перш за все організація обліку всіх форм патології тварин.
Генетичний контроль (моніторинг) шкідливих мутацій повинен включати ретельний клінічний аналіз хвороб і каліцтв, експертизу походження аномальних тварин, з'ясування ролі спадковості в їх етіології. Значення проблеми генетичного моніторингу в сучасному тваринництві пов'язано з багатьма причинами. Так, у зв'язку з використанням штучного осіменіння постійно скорочується число виробників; отже, ступінь впливу кожного з них на генофонд стада, поширення спадкових дефектів значно збільшилася. У зв'язку з цим особливо важливе значення при організації великомасштабної селекції придбала оцінка генотипів биків, кнурів, баранів, які використовуються в інтенсивному відтворенні. Контроль відтворювальних здібностей виробників загальноприйнятими методами за якістю потомства не дає повних відомостей про можливості генетичного впливу їх на запліднюваність, ембріональну смертність, народження аномального і нежиттєздатного, схильного захворювань плода. Ситуація ускладнюється тим, що більшість аномалій і каліцтв - це рецесивно успадковані генні мутації, фенотипично проявляються лише в гомозиготному стані. Успадковані хромосомні аномалії фенотипично проявляються лише у дорослих дочок виробників у вигляді загибелі ембріонів.
Для перевірки виробників на носійство прихованих генетичних дефектів і елімінації їх з відтворення необхідні реєстрація всіх випадків каліцтв і аномалій, контроль стану структури і функцій хромосом.
Організація моніторингу в тваринництві дозволяє контролювати рівні мутагенів у навколишньому середовищі, їх вплив на хромосомний апарат, зростання, розвиток і продуктивність тварин, здійснювати профілактику поширення генетичної патології.
Генетичний вантаж популяцій тварин представлений широкий спектр не тільки генних мутацій, але і аберацій хромосом, які поділяються на кількісні зміни в каріотипі - анеушюідшо (поліплоїдія, гіперплоідія, гіпоплоідія) - і структурні перебудови (транслокація хромосом, інверсії, делеції, нестачі, дуплікації і ін .). Надлишок або нестача хромосом у індивідуума як правило призводить до його загибелі ще на ембріональній стадії розвитку. Виняток становлять носії моносомии, трисомії і деяких інших варіантів анеуплоїдії за статевими хромосомами, які виживають, але є безплідними.
Живі носії структурних перебудов хромосом не мають виражених фенотипичних відхилень. Однак в гаметогенезе у них формуються статеві клітини з незбалансованим набором хромосом, що дають початок нежиттєздатним ембріонам, що служить причиною зниження рівня репродуктивної функції. Ці аберації, будучи збалансованої частиною хромосомних мутацій, передаються у спадок.
Особливості поширення генетичних аномалій тварин. В сучасних умовах розведення тварин, коли генотип виробника за короткий час може бути репродукований тисячами його нащадків, збиток від народження аномального приплоду, зниження його плодючості і життєздатності може бути більше поліпшує ефекту за продуктивністю, якщо виробник є носієм шкідливих генів або аберацій хромосом.
При використанні в розведенні биків, що містять в каріотипі шкідливі гени, а також їх синів і онуків частота генетичної аномалії швидко зростає. Наприклад, в костромський породі інтенсивно використовували бика Бурхана 6083, в потомстві якого було зареєстровано кілька типів каліцтва (вкорочення нижньої щелепи, мопсовідность і витрішкуватість, водянка, сліпота, каліцтва кінцівок і ін.). В результаті, якщо частота цих аномалій в приплоді Бурхана склала 4,87%, то у його онука бика жетони 3501 вже 17,3% нащадків мали каліцтва голови.
Аналіз 14 ліній костромського худоби виявив велике число каліцтв в лінії Ладка. Причиною такого явища зазвичай служить насичення спорідненої групи тварин (лінії) рецесивними мутаціями при відсутності бракування гетерозиготних по шкідливим генам тварин.
Вплив дрейфу генів, концентрація шкідливого аллеля зростають при підвищенні навантаження на одного виробника. Швидкість протікання генетико-автоматичних процесів (дрейфу) залежить від ефективної чисельності популяцій. Для визначення останньої використовують формулу
де Ії - ефективна чисельність популяції; І / - кількість самок; Ат - кількість самців, що беруть участь в розмноженні.
Залежність інтенсивності генетико-автоматичних процесів (дрейфу) від розміру популяції визначається за формулою
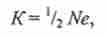
де До - частка, на яку змінюється концентрація алеля.
Так, якщо на поголів'я 1 000 маток будуть використовуватися 5 биків, величина До складе 2,0%, а при використанні одного бика До буде дорівнює 10%. Особливо різко може підвищитися частота мутантного аллеля в популяції, якщо при розведенні лінії генотип гетерозиготного родоначальника буде репродукуватися із застосуванням інбридингу, як це мало місце в лінії Ладка і його продовжувача Бурхана. У п'яти поколіннях цієї лінії зареєстровано 117 телят-виродків. У більшості випадків родоводи батьків і матерів замикалися на трьох предків - биків Бурхана, його батька Ладка і діда Салата. У ряді випадків інбридинг був комплексним (одночасно на зазначених виробників) або посилюється, що підвищувало ймовірність переходу мутантних генів в гомозиготний стан.
При інтенсивному використанні обмеженого контингенту виробників в товарних господарствах генетичну різноманітність популяції звужується навіть при своєчасній ротації ліній. Такий стан може призвести до поєднання родинних по генотипам (гетерозиготних носіїв шкідливих генів) виробників і маток. З іншого боку, інтенсивне кроссирование ліній в племінних господарствах неминуче буде призводити до стихійного родинному парування в товарних господарствах і масового прояву інбредних депресії, в тому числі підвищення частоти каліцтв і аномалій в популяціях.
Інбредних депресія може зрости в результаті міграції. Під міграцією в тваринництві розуміють імпорт виробників, маток або гамет (сперміїв, а також яйцеклітин і ембріонів), закупівлі племінних тварин з інших зон країни. При використанні завезених тварин в місцеву популяцію можуть бути введені не тільки бажані гени, що підвищують продуктивність, а й аллели, що зумовлюють летальні і напівлегальні аномалії. Зрушення концентрацій рецесивного алеля д при міграціях визначаються за формулою
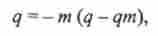
де т - величина обміну генами, а дт - середня концентрація алеля по всій системі популяцій, між якими йде обмін генами, або в тій певної популяції, звідки надходять тварини.
З формули видно, що величина зсуву концентрацій рецесивного алеля залежить від частоти рецесивного алеля в вихідної популяції, звідки мігрують особини, і частоти міграції.
Випадків поширення генетичних аномалій внаслідок міграції відомо чимало. Наприклад, у ФРН в результаті інтенсивного використання на місцевому поголів'я чорно-рябої худоби окремих биків голштинської породи, імпортованих з США, різко зросла частота пупкових гриж у телят. Цю ж аномалію спостерігали в потомстві двох биків голландської породи, закуплених у Чехію з Нідерландів.
Таким чином, щоб не допустити масового поширення спадкової патології, необхідні перевірка генотипів виробників на носійство шкідливих генів і виключення з інтенсивного використання носіїв мутації.
Профілактика поширення летальних і напівлетальних аномалій у сільськогосподарських тварин. Легальні і полулеталь- ні аномалії в основному пов'язані з переходом в гомозиготний стан мутантних рецесивних генів. Це означає, що батьки аномальних тварин є гетерозиготними носіями даних мутацій. Для того щоб шкідливі рецесивні мутації не поширювалися в конкретному стаді або породі, необхідна організація генетичного контролю (моніторингу) за проявом патології у тварин.
Перш за все в кожному стаді повинна бути налагоджена система обліку народження потворного і аномального приплоду. На підставі цього обліку проводиться генетичний аналіз для встановлення ролі спадковості у виникненні аномалій. Такий аналіз повинен підкріплюватися відомостями з літератури щодо етіології даної патології.
Якщо в популяції тварин з'являється вроджена аномалія і встановлено, що вона контролюється аутосомним рецесивним геном, то батьків, від яких отримано аномальний нащадок, і інших фенотипично нормальних їхніх нащадків не залишають для відтворення. Це викликано тим, що батьки є гетерозиготними носіями рецессивной мутації і половина їх нормальних нащадків має такий же ген в гетерозиготному стані. Особливо неприпустимо використання гетерозиготних по шкідливим рецесивним генам виробників в племінних господарствах.
Якщо аномалії виникають у високопродуктивних батьків в племінних стадах, то чоловічих нащадків доцільно перевіряти на гетерозиготное носійство шляхом родинних спаровувань. Для інтенсивного використання виробників надалі відбирають тільки тих з них, у яких при інбридингу не було аномального приплоду. Перевірка виробників за якістю потомства повинна передбачати облік не тільки продуктивності потомства, але і його життєздатності.
Вплив генетичних факторів на мертвонароджуваності і постнатальную смертність молодняка. Мертвонароджуваність і загибель молодняку в перші дні після народження завдають значної шкоди жівотноводсгву. Підраховано, що в загальному доході, що отримується від корови, 10-12% становить дохід від теляти. Мертвими народжуються від 1 до 10% телят. Причини мертвонароджень можуть бути різними. Значну частку (близько 20%) перинатальної смертності приплоду великої рогатої худоби становлять важкі отелення.
На частоту мертвонароджуваності багатодітній родині і вплив генетичні і середовищні чинники, порода і методи розведення батьків.
Встановлено вплив батьків на частоту мертвонароджень і смертності телят після народження. Перинатальна смертність може бути пов'язана із застосуванням інбридингу. Наприклад, в племзаводах «Сичівка», «Нікольський» (Сичевський порода) і «Пролетар» (Костромська порода) відповідно 81,4; 80,0 і 85,8 % мертвих телят народилися в результаті спорідненого спаровування. У цих господарствах було виділено ряд сімейств і пар мати-дочка, де при родинному підборі народжувалися нежиттєздатні телята, а окремі корови неодноразово давали мертвонароджений приплід. У деяких корів після отелення нежиттєздатним телям порушувалася відтворна функція. Вони тривалий час не приносили приплід, що призводило до їх вибракування. Інші корови вибували відразу після отелення мертвим телям.
При перевірці виробників за якістю потомства необхідно вести суворий облік захворюваності і смертності, а також вимушеного забою молодняку. Биков, мають перевагу по ряду селекціоніруемих ознак, але характеризуються підвищеною смертністю нащадків (більше 5%), слід використовувати дуже обмежено.
Для визначення генетичних і середовищних причин смертності приплоду в товарних господарствах необхідно одночасно використовувати сперму не одного, а декількох виробників. В цьому випадку достовірно підвищена частота смертності серед нащадків певного бика за інших рівних умов має розглядатися як прояв генотипу даного виробника.