| Головна |
| «« | ЗМІСТ | »» |
|---|
Одним з основних властивостей матеріалу спадковості є його здатність до самокопірованія - реплікація. Це властивість забезпечується особливостями хімічної організації молекули ДНК, що складається з двох комплементарних ланцюгів. У процесі реплікації на кожній полінуклеотидних ланцюга материнської молекули ДНК синтезується комплементарна їй ланцюг. В результаті з однієї подвійної спіралі ДНК утворюються дві ідентичні подвійні спіралі. Такий спосіб подвоєння молекул, при якому кожна дочірня молекула містить одну материнську і одну знову синтезовану ланцюг, називають напівконсервативним (Див. Рис. 2.12).
Для здійснення реплікації ланцюга материнської ДНК повинні бути відокремлені один від одного, щоб стати матрицями, на яких будуть синтезуватися комплементарні ланцюга дочірніх молекул.
Ініціація реплікації здійснюється в особливих ділянках ДНК, які охоплюють ori (Від англ, origin - початок). Вони включають послідовність, що складається з 300 нуклеотидних пар, впізнавану специфічними білками. Подвійна спіраль ДНК в цих локусах розділяється на два ланцюги, при цьому, як правило, по обидва боки від точки початку реплікації утворюються області розбіжності полінуклео- тідних ланцюгів - Реплікаційний вилки, які рухаються в протилежних від локусу ori напрямках. Між вилки реплікації утворюється структура, звана реплікаціонн'іл оком, де на двох ланцюгах материнської ДНК утворюються нові полінуклео- тідние ланцюга (рис. 3.8, А).
За допомогою ферменту гелікази, що розриває водневі зв'язки, подвійна спіраль ДНК розплітається в точках початку реплікації.
Утворені при цьому одинарні ланцюга ДНК зв'язуються спеціальними дестабілізуючими білками, які розтягують остови ланцюгів, роблячи їх азотисті основи доступними для зв'язування з комплементарними нуклеотидами, що знаходяться в нуклео- плазмі. На кожній з ланцюгів, що утворюються в області реплікаціон- ної вилки, за участю ферменту ДНК-полімерази здійснюється синтез комплементарних ланцюгів (рис. 3.8, Б).
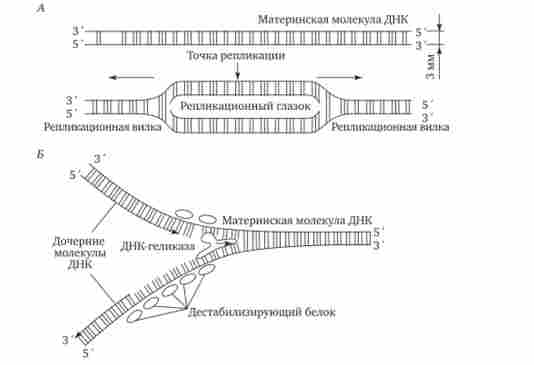
Мал. 3.8. Область початку реплікації. Репликационная вилка.
А - Освіта Реплікаційний вічка;
Б - Область вилки реплікації в молекулі ДНК
У процесі синтезу Реплікаційний вилки рухаються уздовж материнської спіралі в протилежних напрямках, захоплюючи все нові зони.
Поділ спірально закручених ланцюгів батьківської ДНК ферментом геліказу викликає поява супервитки перед репликационной виделкою. Це пояснюється тим, що при розбіжності кожних 10 пар нуклеотидів, що утворюють один виток спіралі, батьківська ДНК повинна зробити один повний оберт навколо своєї осі. Отже, для просування вилки реплікації вся молекула ДНК перед нею повинна була б швидко обертатися, що вимагало б великої затрати енергії. Насправді це не спостерігається завдяки особливому класу білків, званих ДНК-топоізомеразами. Топоізомераза розриває одну з ланцюгів ДНК, що дає їй можливість обертатися навколо другого ланцюга. Це послаблює напругу, що нагромадилася в подвійної спіралі ДНК (рис. 3.9).
До вивільняються водневим зв'язкам нуклеотиднихпослідовностей розділених батьківських ланцюгів приєднуються
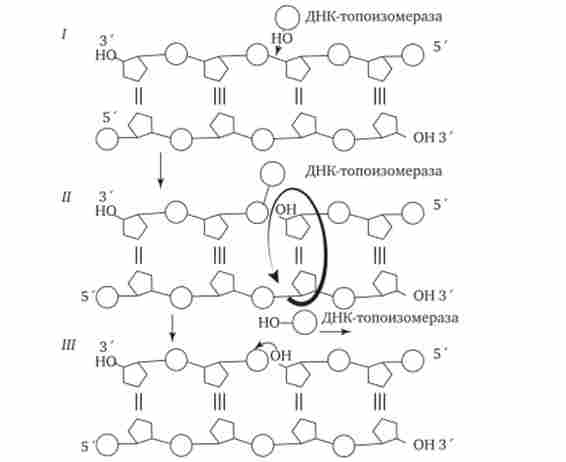
Мал. 3.9. Розрив однієї з ланцюгів ДНК за допомогою ферменту ДНК-топоізомерази.
/ - ДНК-топоізомераза утворює ковалентний зв'язок з однією з фосфатних груп ДНК (верхня ланцюг); II-в результаті розриву фосфодіефірних зв'язку в одній полінуклеотід- ної ланцюга навколо відповідної їй зв'язку інший ланцюга здійснюється обертання, яке знімає напругу, викликане розбіжністю двох ланцюгів ДНК в області репліка- ної вилки; III - після зняття напруги в спіралі ДНК відбувається спонтанне відділення ДНК-топоізомерази і відновлення фосфодіефірних зв'язку в ланцюзі ДНК
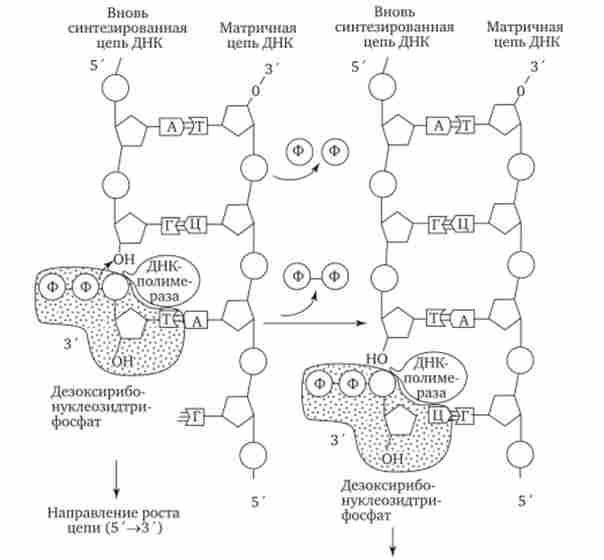
Мал. 3.10. Приєднання чергового нуклеотиду до дочірньої ланцюга ДНК, що синтезується за участю ДНК-полімерази. ФФ - пірофосфат вільні нуклеотиди з нуклеоплазми, де вони присутні у вигляді дезоксирибонуклеозидтрифосфатов: дАТФ, ДГТФ, дЦТФ, дТТФ. Комплементарний нуклеозидтрифосфат утворює водневі зв'язки з певним підставою материнської ланцюга ДНК. Потім за участю ферменту ДНК-полімерази він зв'язується фосфодіефірних зв'язком з попереднім нуклеотидом знову синтезованої ланцюга, віддаючи при цьому неорганічний пірофосфат (рис. 3.10).
Оскільки ДНК-полімераза приєднує черговий нуклеотид до ОН-групі в З'-положенні попереднього нуклеотиду, ланцюг поступово подовжується на її -кінці.
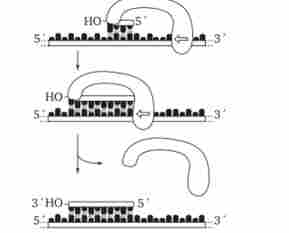
Мал. 3.11. Схема реакції синтезу короткою РНК-затравки, що каталізує РНК-праймазой
Особливістю ДНК-полімерази є її нездатність почати синтез нової полінуклеотидних ланцюга шляхом простого скріплення двох нуклеозидтрифосфатів: необхідний З'-ОН кінець будь-якої полінуклеотидних ланцюга, спареної з матричної ланцюгом ДНК, до якої ДНК-полімераза може лише додавати нові нуклеотиди. Таку полінуклеотидних ланцюг називають затравкой або праймером.
Роль затравки для синтезу полі- нуклеотидних ланцюгів ДНК в ході реплікації виконують короткі послідовності РНК, утворені за участю ферменту РНК-ПРАЙМАЗИ (рис. 3.11). Зазначена особливість ДНК-полімерази означає, що матрицею при реплікації може служити лише ланцюг ДНК, що несе спарену з нею приманку, яка має вільний З'-ОН-кінець.
Здатність ДНК-полімерази здійснювати збірку полинуклеотида в напрямку від 5'до -кінцю при антипаралельними з'єднанні двох ланцюгів ДНК означає, що процес реплікації повинен протікати на них по-різному. Дійсно, якщо на одній з матриць (3 '-> 5') збірка нового ланцюга відбувається безперервно від 5'- до -кінцю і вона поступово подовжується на -кінці, то інша ланцюг, що синтезується на матриці (5 '- > 3 '), повинна була б зростати від 3' до 5'-кінця. Це суперечить напрямку дії ферменту ДНК-полімерази.
В даний час встановлено, що синтез другого ланцюга ДНК здійснюється короткими фрагментами (Фрагменти Окадзакі) Також в напрямку від 5'до -кінцю (по типу шиття «назад голкою»). У прокаріотів фрагменти Окадзакі містять від 1000 до 2000 нуклеотидів, у еукаріот вони значно коротше (від 100 до 200 нуклеотидів). Синтезу кожного такого фрагмента передує утворення РНК-затравки довжиною близько 10 нуклеотидів. Новоутворена фрагмент за допомогою ферменту ДНК-лігази з'єднується з попереднім фрагментом після видалення його РНК-затравки (рис. 3.12, А).
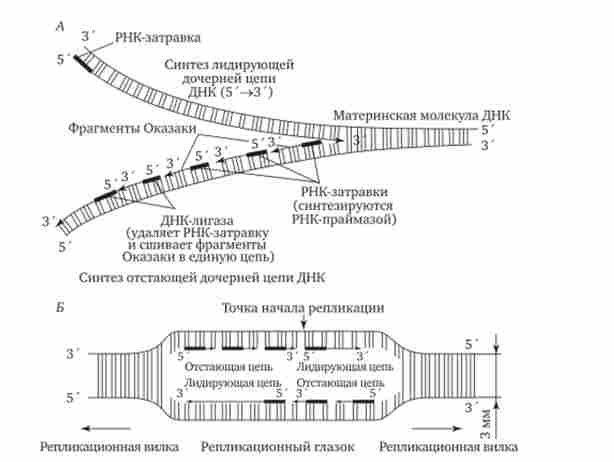
Мал. 3.12. Синтез двох дочірніх ланцюгів ДНК на різних ланцюгах материнської молекули.
А - У зв'язку з антипаралельними ланцюгів ДНК синтез дочірніх ланцюгів йде по-різному: на верхній материнської ланцюга дочірня ланцюг синтезується безперервно - лідируюча ланцюг, на нижній материнської ланцюга дочірня ланцюг збирається з фрагментів Окадзакі - відстає ланцюг; Б - Синтез лідируючих ланцюгів в різноспрямованих вилках відбувається на різних ланцюгах материнської ДНК
У зв'язку з зазначеними особливостями репликационная вилка є асиметричною. З двох синтезованих дочірніх ланцюгів одна будується безперервно, її синтез йде швидше і цей ланцюг називають лідируючої. Синтез інший ланцюга йде повільніше, так як вона збирається з окремих фрагментів, які потребують освіти, а потім видалення РНК-затравки. Тому такий ланцюг називають запізнілої (відстає). Хоча окремі фрагменти утворюються в напрямку 5 '-> 3', в цілому ця ланцюг росте в напрямку 3 ' -> 5 '(рис. 3.12, Л).
З огляду на те що від локусу ori, як правило, починаються дві реплі- Каціон вилки, що йдуть в протилежних напрямках, синтез лідируючих ланцюгів в них йде на різних ланцюгах материнської ДНК (рис. 3.12, Б).
Кінцевим результатом процесу реплікації є утворення двох молекул ДНК, нуклеотидная послідовність яких ідентична такій в материнській подвійної спіралі ДНК.
Розглянута послідовність подій, що відбуваються в ході репликативного синтезу, передбачає участь цілої системи ферментів: гелікази, топоізомерази, дестабілізуючих білків, ДНК-полімерази та інших, спільно діючих в області реплі- Каціон вилки (рис. 3.13).
Реплікація ДНК у про- і еукаріот в основних рисах протікає схоже, проте швидкість синтезу у еукаріот (близько 100 нуклеотидів / с) на порядок нижче, ніж у прокаріот (1000 нуклеотидів / с). Причиною цього може бути освіту ДНК еукаріот досить міцних з'єднань з білками (див. Розд. 3.5.2.), Що ускладнює її деспіраліза- цію, необхідну для здійснення репликативного синтезу.
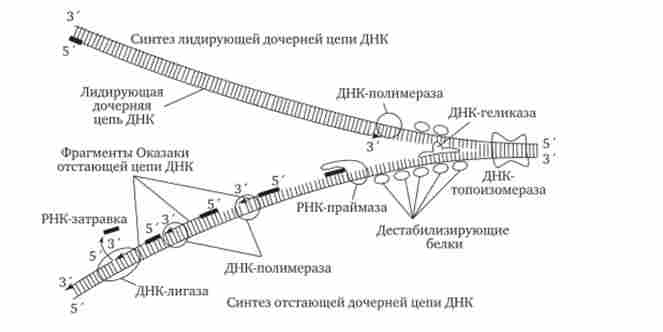
Мал. 3.13. Бедки, які беруть участь в процесі реплікації ДНК.
ДНК-геліказа розплітає подвійну спіраль ДНК, розділяючи її полінуклеотидні ланцюга; дестабілізуючі білки випрямляють ділянку ланцюга ДНК; ДНК-топоізомераза розриває фосфодіефірную зв'язок в одній з полінуклеотидних ланцюгів ДНК, знімаючи напругу, що викликається розплітання спіралі і розбіжністю ланцюгів в вилці реплікації; РНК-праймаза синтезує РНК-затравки для дочірньої ланцюга і для кожного фрагмента Окадзакі; ДНК-полімераза здійснює безперервний синтез лідируючої ланцюга і синтез фрагментів Окадзакі відстає ланцюга; ДНК-лігаза зшиває фрагменти Окадзакі після видалення РНК-затравки
Фрагмент ДНК від точки початку реплікації до точки її закінчення утворює одиницю реплікації - реплікон. Одного разу почавшись в точці початку (локус оп), реплікація продовжується до тих пір, поки весь реплікон НЕ буде дупліціроваться. Кільцеві молекули ДНК прокаріотичних клітин мають один локус ori і являють собою цілком окремі реплікони. Еукаріотичні хромосоми містять велику кількість репліконов. У зв'язку з цим подвоєння молекули ДНК, розташованої уздовж еукаріотичної хромосоми, починається в декількох точках. У різних РЕПЛІКОН подвоєння може йти в різний час або одночасно.