| Головна |
| «« | ЗМІСТ | »» |
|---|
В експериментах з D. melanogaster Т. Х. Морган і його співробітники виявили безліч прикладів зчеплення генів і показали, що це зчеплення як правило неповне.
Розглянемо один з перших експериментів Т. Х. Моргана з вивчення зчепленого успадкування.
У дрозофіли відомі мутантні популяції, що відрізняються від мух дикого типу чорним забарвленням тіла. Це ознака рецесивний по відношенню до ознаки нормальної сірого забарвлення. Ген, який контролює чорне забарвлення тіла, називається black і позначається Ь. Його домінантний аллель - Ь *.
Існує також рецесивний геї vestigial (vg), Який в гомозиготному стані призводить до недорозвинення крил (зародкові крила). Його домінантний аллель (vg+) Контролює нормальний розвиток крил.
При схрещуванні мух $ bb vg vg х b * b * vg * vg ' в / ^ були отримані особини, дігетерозіготние за цими генами. Всі вони були нормальними за обома ознаками відповідно до правила домінування і законом одноманітності F ,. Далі були проведені два типи аналізують схрещувань. У першому з них брали самців F і схрещували з гомозиготними самками bb vg vg, а в другому - незайманих самок, відібраних в F, схрещували з самцями bb vg vg (Рис. 4.1, а). Результати цих аналізують схрещувань виявилися неоднаковими. В F (Потомство від аналізує схрещування) в першому випадку були отримані мухи тільки двох типів незалежно від статі: 50 % мух мали чорне тіло і зародкові крила, 50 % були нормальними за обома ознаками. З огляду на, що розщеплення в анализирующем схрещуванні відображає співвідношення типів гамет, що продукуються особинами F{, слід зробити висновок, що самці F, використані при першому схрещуванні, формували гамети тільки двох типів - з батьківськими поєднаннями алелей b vg і F vg *. Отже, в 100% випадків утворювалися гамети тільки з батьківськими поєднаннями досліджених генів.
При другому схрещуванні в F з'явилися всі можливі чотири типи нащадків, отже, самки Fi давали чотири типи гамет: b vg, b * vg, b vg * і b * vg *. В Fa від цього схрещування чотири типи гамет утворилися в різній кількості (рис. 4.1, б). Незалежно від статі мухи розподілилися наступним чином: 41,5% чорних з зародковими крилами; 41,5 % нормальних за забарвленням і з нормальними крилами; 8,5 % чорних з нормальними крилами; 8,5% нормальних за забарвленням з зародковими крилами. Таким чином, батьківські поєднання b vg і b * vg * утворилися в 83% випадків, а нові комбінації - рекомбінантні поєднання b vg * і b * vg - в 17% випадків.

Мал. 4.2. Батьківські і рекомбінантні поєднання генів
В іншому експерименті в якості батьків були використані мухи, що володіють тими ж ознаками, але в іншому поєднанні: мух з чорним тілом і нормальними крилами схрещували з мухами з нормальною забарвленням тіла і зародковими крилами. Потім знову провели 2 типу аналізують схрещувань: в першому використовували самців Fu у другому - самок F{. В обох випадках їх схрещували з подвійним гомозиготних рецес- сівом bb vg vg, і знову отримали такі ж результати щодо батьківських і рекомбінантних поєднань ознак в Fa. Якщо з брали самців, то спостерігали тільки батьківські комбінації ознак, а якщо з Fx брали самок, то з'являлися батьківські (83%) і рекомбінантні (17%) поєднання ознак в тих же співвідношеннях, що і в першому експерименті.
В обох експерімешдх спостерігається повне зчеплення генів bwvg, якщо для аналізує схрещування беруть самців F. Якщо ж для аналізує схрещування використовують самок F9 то зчеплення часткове.
Т. Х. Морган дав наступне пояснення цих результатів. У першому експерименті гени b і vg знаходяться в одній хромосомі, т. е. дігетерозігот- ні особини F несуть в одному гомологів аллели b і vg, а в іншому гомологів - Ь * і vg*; у другому експерименті b і vg * в одному, а Ь * і vg - в іншому гомологів.
У самців дрозофіли кроссинговер взагалі нс відбувається, тому гени, локалізовані в одній хромосомі (або, кажучи більш строго, в одній парі хромосом), виявляють абсолютне зчеплення, якщо при схрещуванні використовують дігетерозіготних самців. У мейозі у дігетерозігот- них самок дрозофіли F можливий обмін гомологічними ділянками гомологічних хромосом між локусами, в яких знаходяться гени b і vg. Такі обміни, або кроссинговер (Від англ, crossingover - перехрещення), призводять до нового рекомбінантного поєднанню алелей генів b і vg в гомологічних хромосомах, які потім розходяться до різних полюсів. Ці обміни відбуваються з імовірністю 17% і в підсумку дають два класи реці- Прокной рекомбінантних поєднань, або рекомбинантов з однаковою ймовірністю - по 8,5 %.
Подібним чином пояснюється і результат, отриманий раніше У. Бетсоном і Р. Пеннетом: гени, які контролюють забарвлення квіток (р) І форму пилкового зерна (/) у запашного горошку, локалізовані в одній парі гомологічних хромосом, і між ними можливий кросинговер.
Співробітник Т. Х. Моргана А. Стсртсвант припустив, що частота кросинговеру на ділянці між генами, локалізованими в одній хромосомі, може служити мірою відстані, на якому вони знаходяться один від одного. Тоді можна використовувати частоту кросинговеру для того, щоб визначати взаємне розташування генів і відстань між ними.
В якості підтвердження справедливості цього положення можна в загальному вигляді розглянути результати трігібрідного схрещування, в якому батьківські форми дрозофіли розрізняються по вже відомим нам генам b і vg, а також додатково по гену рг, який виявляє зчеплення з Ь і vg. рецесивний аллель pr {purple - пурпурний) в гомозиготному стані обумовлює яскраво-червоне забарвлення очей, нормальна домінантний аллель рг+ - темно-червоний колір очей.
При анализирующем схрещуванні нащадки розщеплюються на 8 класів: 2 класу нерекомбінантні (I і II) і 6 класів нащадків, рекомбінантних по всім генам (Ш-VIII).
Далі необхідно визначити частоту кросинговеру між усіма трьома генами попарно. Для цього підсумовують кількість всіх мух, рекомбінантних по генам ред і рг. класи Ш, IV, VII, VIII. Отримане число ділять на загальне число досліджених нащадків в Fa. Аналогічно визначають частоту рекомбінації (кросинговеру) між рг і vg (При цьому підсумовують класи V, VI, VII, VIII) і частоту рекомбінації між b і vg (підсумовують класи Ш, IV, V, VI).
Експериментально встановлені частоти рекомбінації між трьома генами попарно можна представити таким чином:
На підставі цих даних, користуючись правилом адитивності, можна розташувати 3 гена в лінійній послідовності. Найбільш віддалені один від одного гени b і vg, а між ними локалізовано ген рг. Сума частот його рекомбінації з генами b і vg приблизно дорівнює частоті рекомбінації між Ь і vg. Таким чином, будується найпростіша карта групи зчеплення.
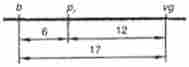
Мал. 4.3. Карта групизчеплення
У строгому сенсі групою зчеплення називають групу генів, які проявляють зчеплене успадкування. Оскільки відомо, що таке спадкування відображає локалізацію генів в одній хромосомі, зазвичай під групою зчеплення розуміють групу генів, розташованих в одній хромосомі.
Гени, розташовані в одній хромосомі, не завжди виявляють зчеплення. Генетічсское відстань, на якому кроссинговер відбувається з ймовірністю I%, вимірюється в сантіморганах (сМ) - ця одиниця виміру, названа на честь Т. Х. Моргана.