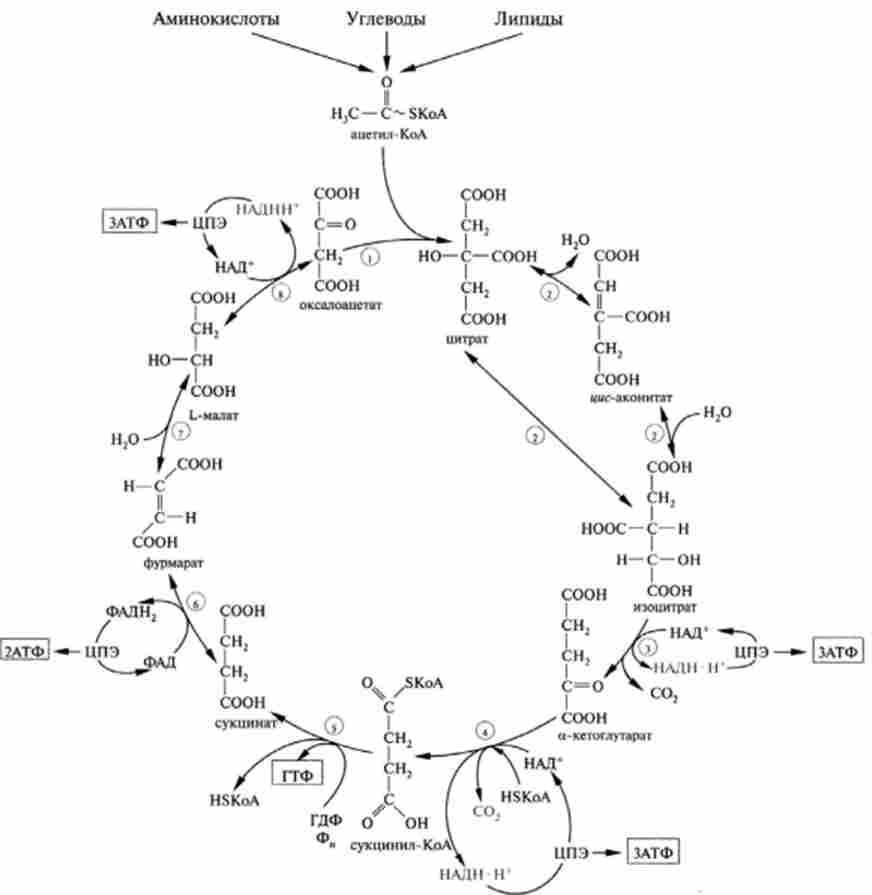
На рис. 19.3 наведена схема реакцій циклу трикарбонових кислот. Як видно зі схеми стехиометричного рівняння ЦТК, в цьому процесі відновлюються три молекули НАДН - Н+ [Реакції (3), (4), (8)] і одна молекула ФАДН2 [Реакція (6) [. Відомо, що при киснево окисленні цих молекул в ланцюзі перенесення електронів в процесі окисного фосфорилювання утворюється при окисленні однієї молекули НАДН - Н+ - ЗАТФ, ФАДН2- 2АТФ. Одна молекула ГТФ (рівнозначно АТФ) утворюється в реакції субстратного фосфорилювання [реакція (5) |.
Мал. 19.3. Схема циклу три карбонових кислот:
ЦПЕ - ланцюг перенесення електронів; в рамках показано число молекул АТФ. утворюються в ЦПЕ в процесі окисного фосфорилювання
Всього це складе: ЗАТФ - 3 + 2АТФ + АТФ = 12АТФ.
Таким чином, за один оборот циклу ТКК утворюється 12 молекул АТФ, з них 11 макроергів - шляхом окисного фосфорилювання і один - на субстратном рівні. Вище на рис. 19.1 наведено розрахунок балансу АТФ при повному аеробному окисленні однієї молекули глюкози (38АТФ), відповідно окислення однієї молекули пірувату складе 15АТФ.
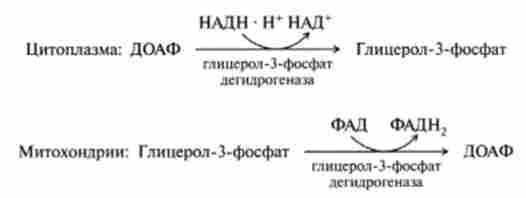
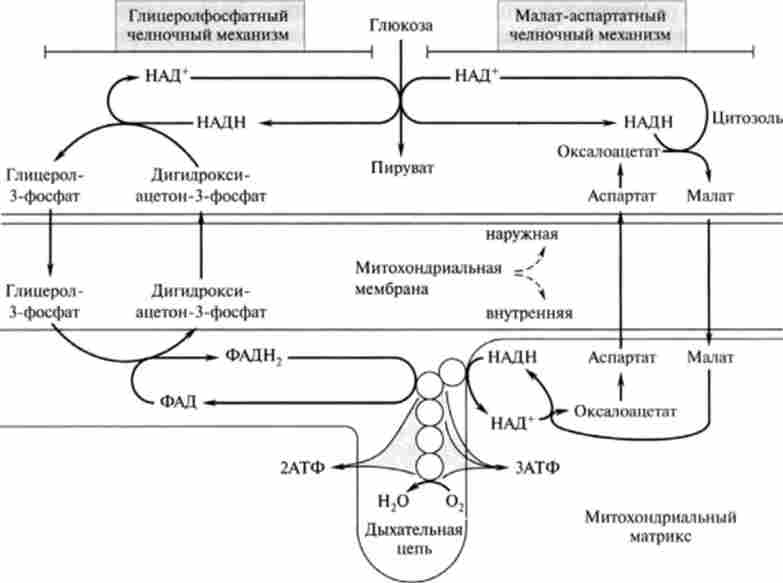
Слід звернути увагу, що відновлені в цитоплазмі в процесі реакції гл і колітічні редукції | гліколіз, реакція (6), гл. 18J дві молекули НАДН можуть при окисленні в мітохондріях давати не шість молекул АТФ, а тільки чотири. Це пояснюється тим, що для НАДН внутрішня мембрана мітохондрій непроникна і вони можуть включатися вдихательная ланцюг
Мал. 19.4. Глінсролфосфатний і малат-аспартатного човникові механізми транспорту НАДН через внутрішню мембрану мітохондрій
за допомогою так званого гл і церола фосфатного човникового механізму (рис. 19.4).
Цитоплазматичний НАДН спочатку відновлює дігідроксіаце- тон-3-фосфат (ДОАФ) до гліцерин-3-фосфату, який легко проникає через мітохондріальну мембрану, де знову окислюється до дігідроксіацстон- 3-фосфату, але при дії ферменту, коферментом якого є ФАД:
Окислення в дихальному ланцюгу ФАДН2 призводить до синтезу НЕ трьох, а двох молекул АТФ. Таким чином, якщо функціонує гл і церола фосфатний човниковий механізм, то при повному окисленні однієї молекули глюкози синтезується 38, а 36 АТФ. Відомо, що за допомогою даного човникового механізму здійснюється перенесення відновлювальних еквівалентів від цитозольного НАДН в мітохондрії в тканинах скелетних м'язів і мозку.
У клітинах печінки, серцевого м'яза і інших функціонує так звана малат-аспартатного човникова система перенесення відновлювальних еквівалентів від цитоплазматичного НАДН в мітохондріальний матрикс. Цей механізм відбувається без витрати енергії, оскільки відновлювальні еквіваленти цитоплазматичного НАДН в мітохондріях відновлюють також НАДН, окислення якого вдихательная ланцюга призводить до синтезу трьох молекул АТФ, і сумарний баланс АТФ при повному окисленні однієї молекули глюкози в цьому випадку складе 38 АТФ.
Біосинтез - біохімія Попередником вітаміну А є ізопснтілпірофосфат, з якого і утворюється молекула каротиноида. Синтез ізопентіл пірофосфату відбувається через синтез мсвалоновой кислоти, яка утворюється в результаті приєднання один до одного трьох молекул ацетил КоА. Мевалонова кислота за допомогою АТФ перетворюється
Біосинтез (анаболізм) рибонуклеотидів - біохімія людини Рибонуклеотиди і дезоксірібо- нуклеотиди є мономерами ДНК, яка знаходиться в хромосомах і мітохондріях і відповідає за зберігання, передачу, трансформацію і реалізацію спадкової інформації. Практично всі живі організми, за винятком деяких видів бактерій, мають здатність синтезувати потрібні
Біологічні мембрани, загальна характеристика, біологічні функції мембран - біохімія Мембранології як самостійна наука, що вивчає будову, властивості, механізми функціонування біологічних мембран, сформувалася порівняно недавно (1950-1970 рр.). Однак сам термін «мембрана» використовується ось уже майже 150 років для позначення клітинної кордону, що служить, з одного боку,
Біологічне значення геномного рівня організації спадкового матеріалу - біологія. Частина 1 Геномний рівень організації спадкового матеріалу, який об'єднує всю сукупність хромосомних генів, є Евола- ционно сформованою структурою, яка характеризується відносно більшою стабільністю, ніж генний і хромосомний рівні. На геномном рівні система збалансованих по дозам і об'єднаних складними
Біокінетики і регулювання біохімічних процесів, введення в біокінетики - біохімія людини Біоенергетика, заснована на законах термодинаміки, дозволяє прогнозувати напрям і глибину мимовільного протікання процесів в залежності від умов (див. Гл. 5). Для цього достатньо розрахувати за допомогою таблиць термодинамічних даних зміна енергії Гіббса AG в процесі. Однак термодинаміка не
Біохімічні основи імунітету, загальна характеристика - біохімія частина 2. Організм людини і тварин містить багато захисних систем і механізмів проти чужорідних речовин і перш за все інфекційних агентів. Мікроби в масі своїй не можуть проникнути в організм завдяки захисній дії шкіри, високій кислотності шлункового соку і ін. Ті чужорідні клітини, які змогли подолати
Білки. Структури і функції, рівні структурної організації білкових макромолекул - біохімія Білки відіграють найважливішу роль в процесах життєдіяльності. Вони є результатом експресії генів і інструментом, за допомогою якого геном управляє всіма метаболічними реакціями в клітці. Білки беруть участь в побудові клітин і тканин, здійснюють біологічний каталіз, регуляторні та скоротливі
Базальні ядра - анатомія центральної нервової системи Базальні ядра включають хвостате ядро, сочевицеподібне ядро, огорожу, мигдалеподібне тіло і прилегле ядро. Найбільшим з цих ядер є хвостате ядро ( п. caudatus). Воно витягнуте в ростри-каудальному напрямку (спереду назад) і має С-подібну форму (рис. 9.1). Мал. 9.1. Базальні ядра: пунктиром