| Головна |
| ЗМІСТ | »» |
|---|
В результаті вивчення даного розділу студенти повинні:
знати
вміти
володіти
Ми вже наголошували, що для збудження і, отже, скорочення м'язового волокна достатньо, щоб до нього прийшов лише один ПД по аксону рухового нейрона. Виникла при цьому деполяризация в мембрані м'язової клітки приводить до генерації «м'язового» ПД, і м'язове волокно скорочується.
Для нейронів, що лежать в межах ЦНС, закономірності збудження виглядають інакше. По-перше, на мембрані однієї нервової клітини є тисячі синапсів, утворених аксонами і дендритами інших нейронів. Деполяризація, викликана активацією одного синапсу, зазвичай мала і не досягає граничної величини, необхідної для збудження постсинаптичного нейрона. Нейрон возбудится тільки в тому випадку, якщо на нього одночасно прийдуть IIД за кількома синаптичним входів від різних нейронів або кілька наступних один за одним з високою частотою ПД по одному і тому ж синапси. По-друге, на сома і дендрі- тах нейрона можуть бути не тільки збуджують синапси, постсинаптична мембрана яких деполяризуется, але і гальмівні синапси, у яких постсинаптична мембрана гиперполяризуется. Гиперполяризация постсінантічної мембрани в більшості випадків досягається шляхом відкриття в ній хлорних каналів, за якими З- по градієнту концентрації спрямовуються в клітину, роблячи потенціал на мембрані ще більш негативним.
Якщо синапс збудливий, то електрод, взнавств в постсинаптичну клітку, зареєструє під дією медіатора місцеву деполяризацію мембрани, яку назвали збудливим постсинаптичним потенціалом (ВПСП) (рис. 9.1, а). ВПСП поширюється електротонічно але мембрані і дуже швидко згасає.
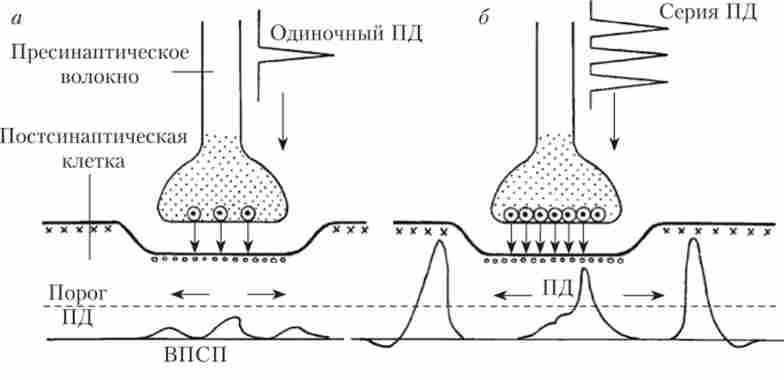
Мал. 9.1. Схема тимчасової сумації:
а - роздратування одного пресинаптичного волокна одиночним імпульсом не приводить до порушення постсинаптичного нейрона, оскільки не досягнуть пороговий потенціал, необхідний для відкриття катіонних каналів в постсинаптичні мембрані; б - серія імпульсів, що приходять по пресинаптичними волокну з високою частотою, викликає більший викид медіатора і сильніший деполяризационного зрушення потенціалу на постсинаптичні мембрані, достатній для генерації ПД постсинаптичної кліткою
Але якщо до синапси прийде серія імпульсів (рис. 9.1, б), То викликані кожним з імпульсів Висі підсумовуються, і зрушення постсинаптичного потенціалу, поширюючись по мембрані, досягає аксонного горбка, де поріг збудження істотно нижче (-60 мВ), і виникає ПД, який почне переміщатися по аксону.
Описуваний варіант синаптичної пластичності називається тимчасової суммацией. Інший варіант - суммация в просторі, або просторова сумація. У цьому випадку на невеликій ділянці мембрани розташовуються синапси, утворені відростками декількох нейронів. Якщо ці синапси одночасно активуються, то виникає сумарного Висі стає досить для того, щоб «спрацювали» потенціалзалежні канали, і в області аксонального горбка виник ПД (рис. 9.2).
З викладеного випливає, що чим ближче синаптічеськоє закінчення розташоване до аксонів горбок, тим більший внесок робить його Висі в запуск ПД, а також чим менше поверхню нейрона, тим швидше і легше
він збуджується, оскільки поширюються по мембрані ВПСП не встигають згаснути.
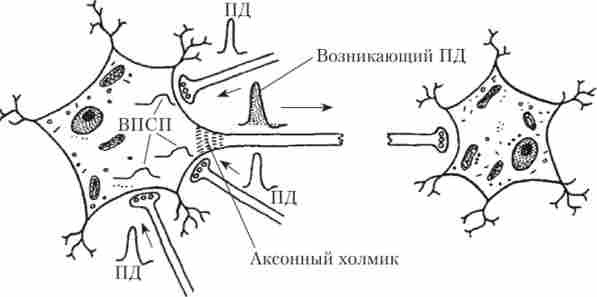
Мал. 9.2. Схема просторової сумації:
ВПСП, що виникає в результаті роботи одного синаптичного входу, недостатньо для того, щоб порушити постсинаптичну клітку. Але сигнали, одночасно прийшли від трьох різних нейронів, підсумовуються і здатні викликати відповідь постсинаптичної клітини у вигляді ПД
Все, що сказано вище, відноситься до збудливим синапсах. Але в ЦНС гальмування представлено не менш, ніж збудження. гальмування являє собою процес придушення збудження, і без гальмування будь-яке порушення неможливо було б послабити або припинити. Порушення може бути місцевим (ВПСП) або поширюється процесом (ПД). Гальмування - локальний процес, пов'язаний з наявністю в нейронних ланцюгах гальмівних нейронів.
Феномен гальмування в ЦНС був відкритий Іваном Михайловичем Сеченовим в 1863 р, коли він виявив, що штучне роздратування структур середнього мозку гальмує у тварин спинномозкові рефлекси.
Процес збудження, що поширюється по нейронних ланцюгів, найпростіше перервати в момент синаптичної передачі, не допустивши проходження нервового імпульсу. При цьому досягти бажаного можна двома шляхами:
У ЦНС виявлені два види гальмування, пресинаптичне і постсинаптичне.
постсинаптичне гальмування зустрічається частіше: на мембрані нейрона, який потрібно загальмувати, розташовуються закінчення гальмівного нейрона, що виділяють гальмові медіатор. Гальмівні медіатори відкривають канати для хлору, вхід якого в клітку через постсинаптичну мембрану генерує гальмівний постсинаптичний потенціал (ТПСП). ТПСП зазвичай являє собою гиперполяризацию постсинаптичної мембрани амплітудою 1-10 мВ і тривалістю близько 10 мс. Таким чином, на короткий час ПП на мембрані в області гальмівного синапсу стане більш негативним. Сусіднім збудливим синапсах в цей момент буде важче порушити гіперполярізованную мембрану, і нейрон буде загальмований.
пресинаптичне гальмування торкається мембрану постсинаптичного нейрона. Воно реалізується за рахунок аксо-аксональні синапсів вставних гальмівних нейронів СМ (рис. 9.3).
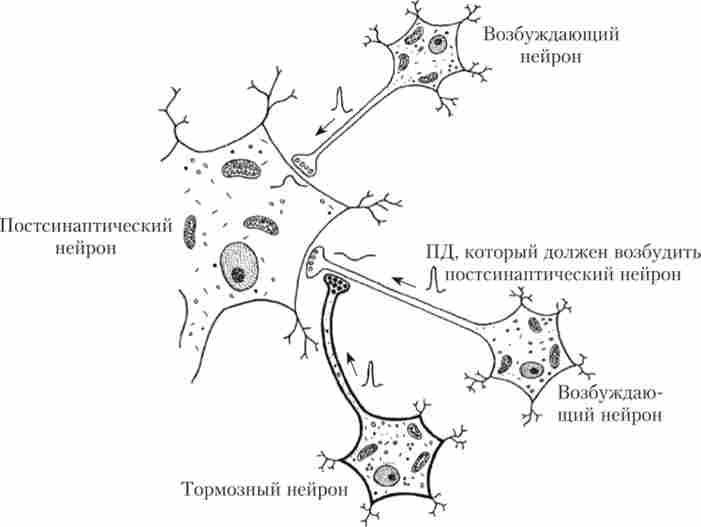
Мал. 93. Схема пресинаптичного гальмування:
стрілки вказують напрям поширення ПД
Механізм пресинаптичного гальмування полягає в тому, що тор- мозний медіатор відкриває хлорні канали в мембрані аксона близько збудливого синапсу, і хлор, виходячи з аксона (там хлорний градієнт відрізняється від такого в сомі), деполяризує його, інактівіруя частина натрієвих каналів і знижуючи амплітуду пресинаптичного ПД. При цьому зменшується надходження кальцію в нервове закінчення, необхідного для синхронного викиду багатьох везикул з збудливим медіатором, - викид збудливого медіатора в синаптичну щілину зменшується або взагалі не відбувається.
Можна виділити дві основні особливості пресинаптичного гальмування. По-перше, воно дозволяє вибірково заблокувати окремий збудливий вхід в клітку. По-друге, ПД, що приходять серіями по аксону гальмівного нейрона, здатні «вивести з ладу» закінчення збудливого нейрона на дуже тривалий за мірками нейрофізіології термін - десяті частки секунди.
Якщо розташовані поблизу один одного збуджуючі і гальмівні синапси активуються практично одночасно, то потенціал на мембрані буде залежати від того, який з цих процесів сильніше. В результаті визначити, виникне чи ні IIД в постсинаптичної клітці в кожен момент, можна за формулою
ПП +? ВПСП _? ТПСП > КУД, де КУД - критичний рівень деполяризації.