| Головна |
| «« | ЗМІСТ | »» |
|---|
В основі м'язового скорочення лежить переміщення ниток актину щодо ниток міозину. Нитки актину рухаються, як по тунелю, між міозіновимі фибриллами, за рахунок утворення зв'язків з міозином. В результаті цього сарко- заходів коротшає (гіпотеза «ковзають ниток» А. Хакслі) (рис. 7.29). При цьому довжина 1-дисків зменшується, А-диски зберігають свій розмір.
Ковзання Актинові і міозінових ниток один щодо одного можливо тільки в присутності іонів Са2+ і АТФ, яка утворюється при розщепленні глікогену, глюкози і жирних кислот. М'язи характеризуються активним обміном речовин. До них підходить велика кількість кровоносних і лімфатичних судин, а також нервів. Останні утворюють з м'язовими волокнами синаптичні контакти.
Всю ланцюг подій при м'язовому скороченні можна представити таким чином: в нервово-м'язовому синапсі під впливом приходять з ЦНС але нервового волокна імпульсів виділяється медіатор ацетилхолін, Деполяризуючий мембрану м'язового волокна. Виникає при цьому імпульс поширюється по мембрані волокна і Т-трубочках і передається на мембрану саркоплазматичного ретикулума, з якого в саркоплазму виходить кальцій. Іони кальцію сприяють утворенню комплексу акто- міозину і розщепленню АТФ; звільняється при цьому, забезпечує ковзання тонких Актинові ниток вздовж міозінових.
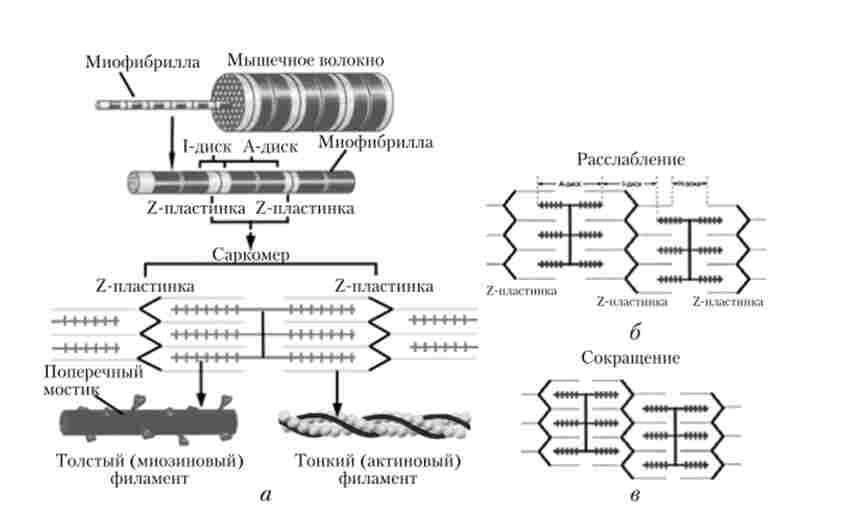
Мал. 7.29. Будова міофібрили (а).
Зміна взаємного розташування міофібрил при розслабленні (б) і скорочення (в) м'язового волокна
Розслаблення м'яза пов'язане зі зворотним надходженням Са2+ в саркоплазматичний ретикулум, що відбувається за участю активних механізмів, пов'язаних з роботою іонних насосів. Якщо концентрація іонів кальцію в саркоплазме знижується і вони перекачуються в ендоплазматичну мережу, то скорочення м'язового волокна припиняється.
Скелетні м'язи людини складається з м'язових волокон декількох типів з різними структурно-функціональними характеристиками. Виділяють чотири основних типи м'язових волокон: повільні фазічні волокна окисного типу, швидкі фазічні волокна окисного типу, швидкі фазічні окисні волокна з гліколітичні типом окислення і тонічні волокна.
Повільні фазічні м'язові волокна окисного типу містять велику кількість білка міоглобіну, який зв'язує 02. Цей білок аналогічний гемоглобіну еритроцитів і надає м'язових волокон темно-червоне забарвлення. М'язи, що складаються переважно з цих волокон, беруть участь в підтримці пози людини. Втома в них розвивається дуже повільно, а відновлюються функції дуже швидко.
М'язи, що складаються переважно з швидких фазі- чеських волокон окисного типу, виконують швидкі сокрашенія без помітного стомлення. Це пов'язано з наявністю в волокнах великої кількості мітохондрій і хорошою здатністю синтезувати АТФ. Основне призначення таких волокон - виконання швидких, енергійних рухів.
М'язи, що містять швидкі фазічні волокна з гли- колитичним типом окислення, розвивають швидке і сильне скорочення, але порівняно швидко втомлюються. АТФ в волокнах цього типу утворюється за рахунок гліколізу. Міоглобін в них відсутня, тому їх називають «білими волокнами».
тонічні волокна скорочуються і розслабляються повільно, так як активність АТФ в них низька. Такі волокна входять до складу деяких м'язів очі.
Більшість скелетних м'язів людини складається з м'язових волокон різних типів з переважанням одного з них в залежності від функцій, які виконує та чи інша м'яз.
Основне фізіологічне властивість м'язів - скоротність - проявляється в здатності м'язи до укорочення або розвитку напруги. Розрізняють два типи м'язових скорочень - ізотонічні і ізометричні. при фізіологічному скороченні волокна м'язи коротшають, але напруга залишається постійним. при изометричному - м'яз вкоротити не може, довжина м'язових волокон залишається незмінною, так як обидва се кінця нерухомо закріплені, але напруга в міру їх скорочення зростає.
По відношенню до цілого організму застосовується інша класифікація типів скорочення: изометричним називають скорочення, при якому довжина м'яза не змінюється, концентричних - при якому м'яз коротшає, ексцентричним - подовжується (наприклад, при повільному опусканні вантажу). Для природних рухів зазвичай характерні всі три типи скорочення м'язів.
Функціональною одиницею скелетної мускулатури вважається не окреме м'язове волокно, а нейромоторная, або рухова одиниця, яка включає кілька м'язових волокон, що іннервуються мотонейроном спинного мозку (рис. 7.30, 7.31). У відповідь на імпульси, що приходять від мотонейрона, скорочуються всі м'язові волокна, що входять до нейромоторную одиницю.
Число м'язових волокон, складових рухову одиницю, швидкість їх скорочення і стійкість до стомлення неоднакові. Залежно від їх властивостей рухові одиниці поділяють на швидкі (фазні)} повільні (тоніРіс. 7.30. рухові одиниці
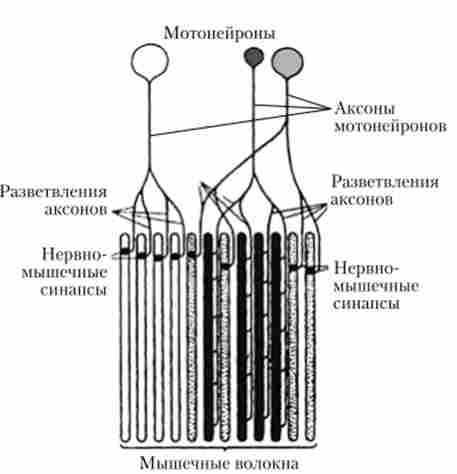
чноё) і перехідні. Рухові одиниці кожного м'яза неоднакові. М'язи, що забезпечують точні і швидкі рухи (наприклад, м'язи пальців руки), складаються в основному з декількох сотень або тисяч швидких рухових одиниць. У більшості м'язів змішаного типу першими активізуються повільні рухові одиниці, розвиваючі
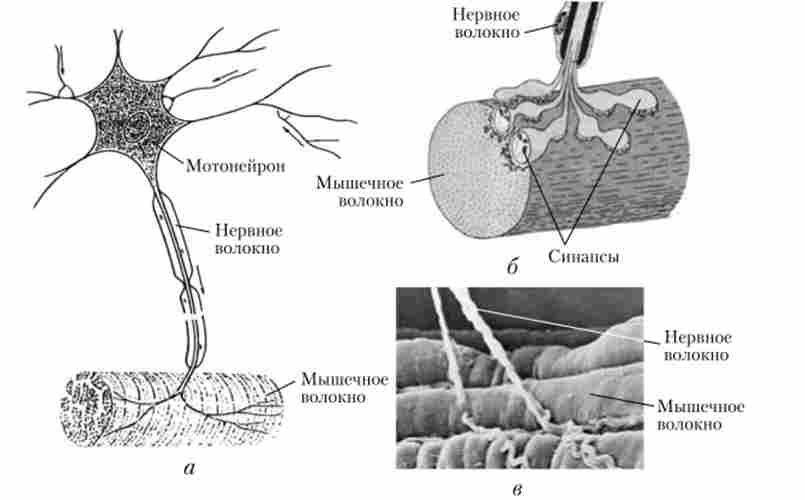
Мал. 7.31. Нервово-м'язове з'єднання:
а, 6 - нервово-м'язовий синапс; в - електронна скануюча
мікроскопія невелику силу скорочення, а при збільшенні збудження в скорочення залучаються м'язові волокна, розвиваючі велику силу. Активація швидких нсйромогорних одиниць забезпечує точну рухову реакцію.
У природних умовах до м'яза з ЦНС поступають не поодинокі імпульси, а серії імпульсів, на які вона реагує не поодиноким, а тривалим (тетанічних) Скороченням. Вона зумовлена тим, що кожен наступний імпульс приходить в момент, коли ще не закінчилася попередня хвиля скорочення. Остання, підсумовуючись з попередньою, продовжує скорочення м'язи. Якщо кожна нова хвиля скорочення виникає в момент, коли м'яз вже почала розслаблятися під впливом попереднього роздратування, виникає зубчастий тетанус. При меншому інтервалі між подразненнями, коли кожна нова хвиля скорочення виникає до початку м'язового розслаблення, утворюється суцільний, або гладкий, тетанус. Окремі волокна м'язи при її природної стимуляції з нерва відповідають на кожен імпульс одиночним скороченням. Злитий тетанус виходить за рахунок сумації скорочень окремих м'язових волокон. Зазвичай м'язові волокна одного м'яза скорочуються не одночасно, так як імпульси від різних мотонейронів ЦНС також приходять до них не одночасно. Це сприяє утворенню та підтриманню злитого тетанічес- кого скорочення м'язи.
Скорочуючи, м'яз виконує роботу. Робота м'язів залежить від сили їх скорочення, а сила скорочення однієї і тієї ж м'язи - від кількості нейромоторную одиниць, що беруть участь в ньому. Чим їх більше, тим скорочення інтенсивніше. Сила скорочення також залежить від частоти подразнення. До певної межі збільшення частоти стимуляції супроводжується зростанням сили м'язового скорочення. Це пов'язано з тим, що зі збільшенням частоти роздратування в реакцію включається все більша кількість м'язових волокон. Максимальна напруга, яке може розвинути м'яз, визначається числом волокон, з: чим воно більше, тим більше сила м'язів. У зв'язку з цим перисті м'язи, що складаються з безлічі волокон, відрізняються більшою силою.
Прояв сили м'язи залежить і від особливостей її прикріплення до кісток. М'язи з більшою площею прикріплення або опори, мають великі можливості для прояву сили. Важливо також і місце прикладання сили м'язів. Кістки разом з прикріпляються до них м'язами є важелями, тому чим ближче до точки докладання сили тяжіння або чим далі від точки опори важеля і ближче до точки докладання сили тяжіння прикріплюється м'яз, тим більшу силу вона може розвинути (рис. 7.32).
Залежність м'язової сили від таких факторів чітко проявляється в діяльності м'язів верхньої та нижньої кінцівок. Верхня кінцівка призначена для виконання різноманітних точних і швидких рухів. Функція нижніх кінцівок вимагає великої сили їх м'язів. Цим функціональним завданням відповідає і характер прикріплення відповідних м'язів. Так, дельтоподібний м'яз, розташована в області плечового суглоба, має невелику поверхню опори і прикріплюється на плечовій кістці поблизу від місця опори важеля. У м'язів нижніх кінцівок площу опори велика і точка прикладання сили знаходиться далеко від точки опори. У сідничного м'яза площа опори в 23 рази більше, ніж у дельтоподібного, а площа прикріплення - більше в 4,5 рази.
Між силою м'язи і величиною її укорочення немає прямо пропорційної залежності. Максимальна вкорочення м'яза, а отже, і інтенсивність скорочення, що викликається цим укороченням руху, в тому чи іншому суглобі залежить від довжини м'язових волокон. Вона найбільша в м'язах з паралельним розташуванням волокон, в той час як більшою силою володіють перисті м'язи. Спочатку розтягнута м'яз при скороченні коротшає на велику величину.
Робота м'язи при скороченні дорівнює добутку маси вантажу на підняту висоту. Звідси випливає, що максимальна робота, виконувана при одиночному скороченні м'яза, залежить від її сили (чим більше сила, тим більший вантаж може бути піднятий) і ступеня укорочення м'яза. У про-
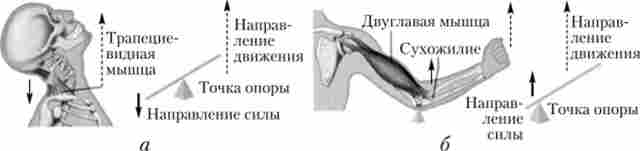
Мал. 732. Важелі тіла людини:
а - важіль рівноваги; б - важіль швидкості. Трикутник - точка опори; темні стрілки показують напрямок сил м'язової тяги; пунктирні стрілки - напрям сили тяжіння; пунктирна стрілка - рух процесі природної діяльності людини величина роботи, що виконується тією чи іншою м'язом, в значній мірі залежить від се здатності тривалий час перебувати в скороченому стані (витривалість м'язів). Розрізняють витривалість до статичних і динамічних зусиль. Витривалість до статичних зусиль визначається часом, протягом якого утримується величина заданого зусилля. У різних м'язів вона неоднакова. Найменшою витривалістю характеризується триголовий м'яз плеча (1 хв - при зусиллі, що дорівнює 50% максимального), найбільшою - литковий м'яз (7 хв).
Витривалість до тривалої роботи залежить не тільки від величини вантажу, що піднімається, але і від темпу роботи. Робота буває найбільшою при якійсь середній величині вантажу і частоті рухів. Для кожного виду м'язової діяльності можна підібрати певний середній (оптимальний) ритм і величину навантаження, при якій робота стане максимальною, а втома розвиватиметься поступово.
Робота м'язів - необхідна умова їх скорочення. Тривала бездіяльність веде до атрофії м'язів і втрати працездатності. Помірна систематична робота м'язів сприяє збільшенню їх обсягу, зростанню сили і працездатності, що важливо для фізичного розвитку всього організму.
При тривалій динамічної або статичної роботі настає стомлення м'язів. втомою називають тимчасове зниження працездатності клітини, органу або цілісного організму, що наступає в результаті роботи і зникає після відпочинку. У природних умовах стомлення пов'язано, перш за все, зі змінами, що відбуваються в нервовій системі, зокрема з порушенням проведення збудження в межнейрональних і нервово-м'язових синаптичних контактах. Швидкість настання втоми залежить від стану нервової системи, ритму, в якому проводиться робота, і від величини навантаження. Після відпочинку працездатність відновлюється. І. М. Сєченов вперше (в 1903 р) показав, що відновлення працездатності стомлених м'язів руки людини після тривалої роботи з підйому вантажу відбувається швидше, якщо в період відпочинку виконувати роботу іншою рукою або ногою. Такий відпочинок був названий активним.
Чергування розумової та фізичної праці, динамічні паузи до і під час занять сприяють підвищенню працездатності дітей і дорослих. Чим менша дитина, тим швидше у нього розвивається стомлення. У грудному віці стомлення настає через 1,5-2 години звичайного неспання. Діти стомлюються і при нерухомості або тривалому обмеження рухів.
М'язи людини навіть в спокої знаходяться в дещо скороченому стані. Тривале утримання напруги називають м'язовим тонусом. Під час сну або при наркозі тонус м'язів знижується і внаслідок цього тіло розслабляється. Тонічні скорочення м'язів не призводять до розвитку втоми. Повне зникнення тонусу м'язів спостерігається тільки після смерті. Збереження тонусу обумовлено постійним надходженням до м'яза наступних один за одним з великими інтервалами нервових імпульсів від рухових нейронів Ц11С. Активність цих нейронів підтримується імпульсами, які надходять з верхніх відділів ЦНС і від рецепторів м'язів - м'язових веретен.
Тонус м'язів грає важливу роль в здійсненні координації рухів. У новонароджених і грудних дітей переважає тонус м'язів-згиначів, обумовлений підвищеною збудливістю червоного ядра середнього мозку. У міру функціонального дозрівання пірамідної системи мозку і нейронів кори великих півкуль тонус м'язів у дітей знижується. Це чітко проявляється в другому півріччі життя дитини і є необхідною умовою для розвитку ходьби. До трьох - п'яти років встановлюється рівновага тонусу м'язів-аітагоністов.