| Головна |
| «« | ЗМІСТ | »» |
|---|
Перетворення глюкози в пировиноградную кислоту, що каталізує ферментами гліколізу, є центральним шляхом катаболізму вуглеводів в більшості клітин як в аеробних, так і в анаеробних умовах (див. Розд. 9.1). Точно так же зворотний процес перетворення пірувату в глюкозу є центральним шляхом біосинтезу моносахаридів і полісахаридів.
У центральний шлях глюконеогенезу вливаються два шляхи, що починаються з різних наборів невуглеводних попередників.
Один з цих шляхів складається з ряду послідовних реакцій, в яких проміжні продукти циклу трикарбонових кислот перетворюються в пировиноградную кислоту.
Другий шлях складається з ряду реакцій, що призводять до відновлення С02 до глюкози. Цей шлях відсутній у гетеротрофних організмів і є відмінною рисою автотрофов, особливо фотосинтезирующих клітин.
Розходяться шляхи, які починаються з глюкозо-6-фосфату (див. Рис. 9.9) призводять до утворення вільної глюкози, запасних полімерів (крохмалю і глікогену) та інших моносахаридів і їх похідних.
Ще один важливий шлях глюконеогенезу починається з амінокислот. Всі організми можуть перетворювати амінокислоти в різні проміжні продукти циклу трикарбонових кислот і потім в глюкозу. Однак кількості утвореної
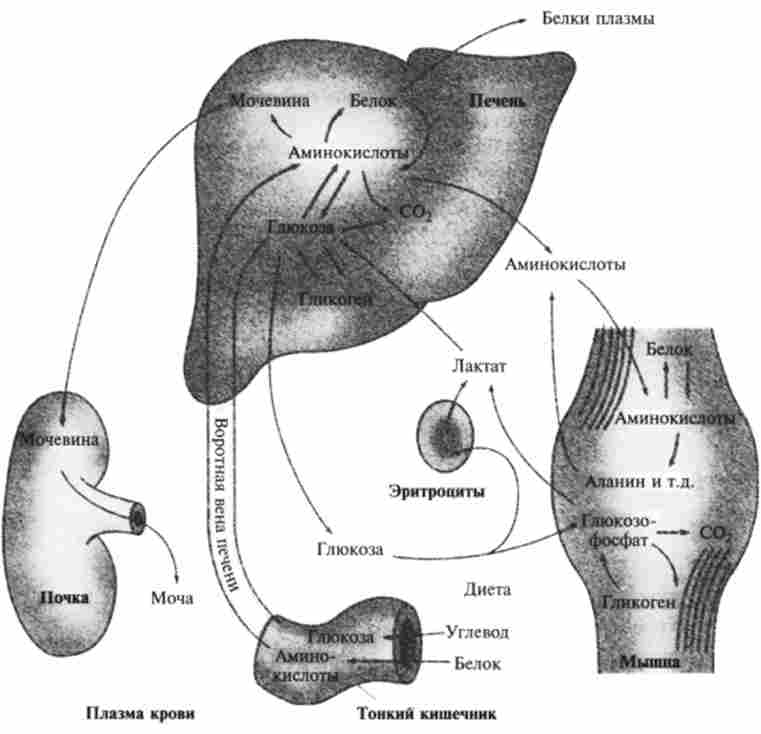
Мал. 9. / 0. Глюкоза і ес метаболіти в процесах життєдіяльності
при цьому глюкози можуть значно варіювати в залежності від потреби організму в амінокислотах для синтезу білка і від доступності інших видів клітинного палива.
У людини і вищих тварин в період відновлення після інтенсивного м'язової роботи особливо активно відбувається синтез глюкози з молочної кислоти (лактату) крові, який наразі триває головним чином в печінці.
Шляхи, що ведуть від глюкозо-6-фосфату до інших продуктів, розрізняються у різних організмів. Здатність до утворення вільної глюкози порівняно обмежена; нею володіють лише деякі рослини, а також клітини печінки, нирок і тонкого кишечника хребетних. У той же час шляхи, що ведуть до синтезу крохмалю і глікогену, мабуть, майже універсальні, але використовуються в різному ступені в залежності від потреб обміну та ресурсів харчування. Слід зазначити, що шляхи, що ведуть до утворення позаклітинних глікогену і крохмалю, сильно диференційовані і відрізняються високою специфічністю.
Центральний шлях, що веде від пірувату до глюкози, - освіту фосфо енолпірувата з пірувату. Більшість реакцій биосинтетичного шляху утворення глюкози з пірувату каталізується ферментами гликолитичного циклу. Таким чином, ці реакції протилежні реакціям, що реалізується в процесі гліколізу (див. Рис. 9.2). Однак в нормальному гл і колітічні шляху, т. Е. В шляху «вниз», є три стадії, які не можуть використовуватися при перетворенні пірувату в глюкозу, т. Е. В шляху «вгору».
Глюконеогенез йде в обхід необоротних стадій, використовуючи альтернативні реакції, які термодинамічно вигідні. Перша з них - перетворення пірувату в фосфоенолпіруват:
Ця реакція не йде шляхом прямого звернення бенкеті ватки скарбницею реакції внаслідок великого позитивного зміни стандартної енергії Гіббса (ДG ° ' = +30 кДж / моль).
Фосфорилювання пірувату досягається обхідним шляхом послідовності реакцій, каталізуються як ферментами цитоплазми, так і ферментами мітохондрій. В результаті численних досліджень було встановлено ферменти цієї послідовності.
У першій реакції ферментом є мітохондріальна піруваткарбоксіла- за, яка каталізує реакцію утворення проміжних продуктів циклу трикарбонових кислот з пірувату:
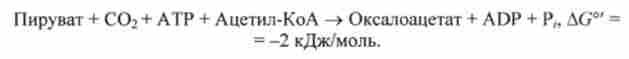
Піруваткарбоксілази - регуляторний фермент, який неактивний відсутність ацетил-КоА. Оксалоацетат, що утворюється в цій реакції, відновлюється потім в мітохондріях в малат - реакція, що йде за участю NADH:
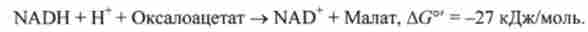
Потім малат дифундує з мітохондрій в цитоплазму, де він окислюється цитоплазматичної NAD-залежної формою малатдегідрогенази з утворенням внемітохондріального оксапоацетата:
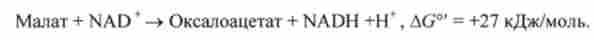
Хоча, як видно з рівняння, ця реакція є сильно ендергонічеськие, проте її рівновагу зрушено вправо, оскільки її кінцевий продукт швидко віддаляється. Під дією фосфопіруваткарбоксілази з оксапоацетата утворюється фосфоенолпіруват. Донором фосфату в цій реакції служить GTP:
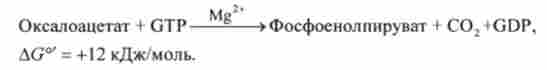
Цей фермент виявлений в цитоплазмі клітин печінки пацюки та миші, в мітохондріях кролика і курчати, а також в цитоплазмі і мітохондріях морської свинки.
Сумарне рівняння для цього обхідного шляху освіти фосфоенолпіру- вата отримують, підсумовуючи все реакції і зміни енергії Гіббса:
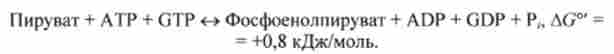
Ця сумарна реакція оборотна, оскільки загальна зміна стандартної енергії Гіббса невелика. При високому відношенні концентрацій [АТР] / [ADP] і в присутності надлишку пірувату рівновагу буде зрушено вправо.
Можна розділити цей процес на ендергонічеськие і екзергонічеськие реакції.
Ендергонічеськие (що йде зі споживанням енергії) реакцією є перетворення пірувату в фосфоенолпіруват:
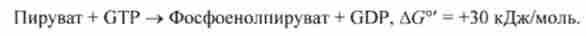
Гідроліз АТР є екзергонічеськие реакцію:
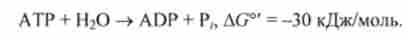
Ясно, що на фосфорилирование однієї молекули пірувату витрачається в кінцевому рахунку енергія двох високоенергетичних фосфатних зв'язків АТР і GTP, кожній з яких відповідає величина AG ° рівна -30 кДж / моль.
Обхідний шлях, що веде до утворення фосфоенолпіруват, заснований на тому, що в мітохондріях відношення концентрацій NADH / NAD 'відносно велике. Тому мітохондріальний оксалоацетат легко відновлюється в малат.
У цитоплазмі відношення концентрацій NADH / NAD 'дуже мало і поза мітохондрій малат знову окислюється в оксалоацетат. Малат, легко проходить через мітохондріальну мембрану, служить переносником відновлювальних еквівалентів між двома відсіками клітини.
Фосфоенолпіруват, утворений з пірувату в результаті описаних вище реакцій, легко перетворюється в фруктозо-1,6-дифосфат в послідовності реакцій, які представляють собою реакції гліколізу, що йдуть у зворотному напрямку.
Вільний гліцерин, утворений при гідролізі жирів - тріацілгліцері- дов, і гліцерил-3-фосфат служать попередниками глюкози. Вони включаються в послідовність реакцій, що ведуть від пірувату до глюкози, після перетворення в діоксіацстонфосфат.
Реакція освіти фруктозо-6-фосфату не йде в напрямку синтезу глюкози:
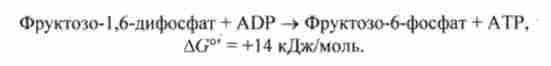
Біосинтез глюкози йде в обхід цієї ендергонічеськие реакції за допомогою ферменту гексозодіфосфатази. Цей фермент здійснює незворотний гідроліз 1,6-дифосфатні групи:
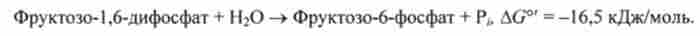
Встановлено, що гексозодіфосфатаза є регуляторний фермент, активність якого сильно пригнічується АТР. Цей фермент має три або більше центру зв'язування АМР. Фермент основну активність виявляє в напрямку синтезу глюкози, коли концентрація АМР мала, а концентрація АТР велика.
Наступного (оборотною) стадії біосинтезу глюкози фруктозо-6-фосфат перетворюється в глюкозо-6-фосфат за участю фосфогексозоізомерази:
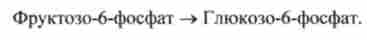
У більшості клітин глюкозо-6-фосфат, що утворюється в процесі глюко- неогенеза, використовується як попередник запасних полімерів, різних моносахаридів (крім глюкози), дисахаридів і структурних полімерів.
У клітинах ряду органів, наприклад печінки, нирок і кишкового епітелію, хребетних глюкозо-6-фосфат може отщепить фосфорну групу (дефосфорілі- роваться) з утворенням вільної глюкози.
Вільна глюкоза виходить при дії глюкозо-6-фосфатази, що каталізує кінцеву реакцію глюконеогенезу:
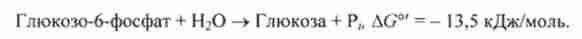
У кров синтезована глюкоза надходить головним чином з печінки. Глю- козо-6-фосфатаза не міститься в м'язах і в мозку, відповідно з цих тканин вільна глюкоза в кров не надходить.
Сумарне рівняння метаболічного шляху глюконеогенезу, який веде з пи- рувата до вільної глюкози, має вигляд
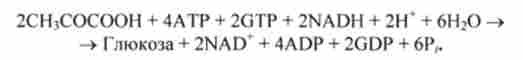
На освіту кожної молекули глюкози витрачається шість високоенергетичних фосфатних зв'язків АТР і GTP, і дві молекули NADH використовуються в якості відновників.
Сумарна реакція протікає з виділенням енергії. Легко бачити, що це рівняння сильно відрізняється від рівняння зворотної реакції перетворення глюкози в пировиноградную кислоту, яка також є екзергонічеськие:
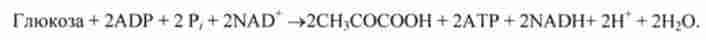
Синтез глюкози з пірувату контролюється вмістом «палива» для дихання (ацетил-КоА), а також вмістом енергетичного заряду системи АТР. Синтез глюкози прискорюється при утворенні надлишкової кількості мітохондріального ацетил-КоА, що перевищує ту кількість, яка клітина може використовувати в даний момент в якості палива.
Описаний вище шлях синтезу глюкози з пірувату відкриває також можливість для утворення глюкози з різних попередників пірувату або фосфоенолпіруват. Головними з них є проміжні продукти циклу трикарбонових кислот, які можуть окислюватися в малат. Потім малат може виходити з мітохондрій і окислюватися в цитоплазмі в оксалоацетат з подальшим утворенням фосфоенолпіруват під дією цитоплазматичної фосфо енолпіруваткарбоксікінази.
У тканинах тварин є альтернативний шлях утворення фосфоенолпіруват з а-кетоглутарата і попередніх йому 6-вуглецевих кислот циклу Кребса.
В кінцевому рахунку при будь-якій послідовності реакцій три атома вуглецю різних проміжних продуктів циклу трикарбонових кислот перетворюються в три атома вуглецю фосфопірувата.
За допомогою різних експериментів встановлено також, що у хребетних проміжні продукти циклу трикарбонових кислот використовуються для глюконеогенезу. Наприклад, в одній серії дослідів щури голодували 24 год або більше. При цьому рівень глікогену в печінці знизився приблизно від 7 до 1% сирої маси. Подальше згодовування бурштинової кислоти або інших проміжних продуктів циклу трикарбонових кислот голодуючим щурам викликало збільшення загальної кількості глікогену, головним чином завдяки підвищенню рівня глікогену в печінці.
Перетворення проміжних продуктів циклу трикарбонових кислот в глюкозу спостерігали також у тварин, які отримували токсичний глікозид флоридзин. Флоридзин блокує реабсорбцію глюкози з ниркових канальців, внаслідок чого глюкоза крові майже повністю виводиться з сечею. Виявилося, що згодовування бурштинової кислоти або інших проміжних продуктів циклу трикарбонових кислот тваринам, які мають флоридзин, призводить до виділення глюкози в кількостях, майже точно еквівалентних змістом вуглецю в цих проміжних продуктах, отриманих тваринами.
У тканинах вищих тварин істинний синтез глюкози з двох атомів вуглецю ацетильной групи ацетил-КоА не відбувається. Одна з причин цього полягає в тому, що цитрат - шестіуглеродний продукції кт конденсації ацетил-КоА і ок- салоацетата - в циклі Кребса при окисленні в фосфоенолпіруват втрачає три атома вуглецю у вигляді СО2. Отже, цитрат не може служити джерелом додаткових кількостей глюкози понад те, що утворюється з оксалоацетата. Крім того, в тварин тканинах ацетил-КоА не може безпосередньо перетворюватися ні в піруват. ні в сукцинат. У вищих тварин метаболічний шлях, в якому атоми вуглецю жирних кислот могли б використовуватися для глюконеогенезу, Відсутнє.
Навпаки, рослини і багато мікроорганізмів можуть синтезувати вуглеводи з жирних кислот через ацетил-КоА за допомогою реакцій глиоксилатного циклу. Для такого перетворення необхідні два специфічних ферменту - ізоцітратліа- за і малатсінтаза. Обидва ці ферменту відсутні у вищих тварин.
Сукцинат, що утворюється в глиоксилатного циклі, перетворюється в оксапоаце- тат, який, в свою чергу, служить попередником фосфоенолпіруват. Саме за допомогою цього шляху запасні жири перетворюються в глюкозу в проростають насінні.