| Головна |
| «« | ЗМІСТ | »» |
|---|
Всі відомі механорецептори за своїми функціональними характеристиками можна розділити на кілька груп. В основі класифікації механорецепторов лежить їхня реакція на ступенчатообразние механічні стимули, у яких змінюється амплітуда, швидкість її наростання і спаду, тривалість і частота. Крім ступенчатообразних стимулів широко застосовуються синусоїдальні механічні стимули. Механорецептори, що входять до складу будь-якої групи, налаштовані на сприйняття певного параметра механічного стимулу.
У шкірі виявлені механорецептори, за своїми функціональними характеристиками подібні до м'язовими веретенами, т. Е. Детектори амплітуди - швидкості (рис. 15.6, А). У спокої ці рецептори мають спонтанної активністю. У відповідь на постійне
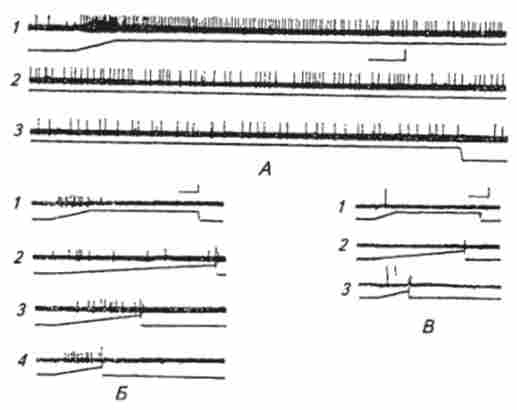
Мал. 15.6. Електричні відповіді шкірних механорецепторів на механічні стимули різної форми:
А. Імпульсна активність медлснноадаптіруюшегося рецептора (детектора амплітуди-швидкості) на ступенчатообразний механічний стимул тривалістю близько 10 з: / - початок; 2-продовження; 3 закінчення електричного відповіді рецептора; калібрування по вертикалі 0,5 мВ, по горизонталі 250 мс. Б. Імпульсна активність сь- роадаптіруюшіхся мсханорсіспторов (детекторів швидкості) на ступенчатообразний (/) і пилковидні (2, 3, 4) механічні стимули з різною швидкістю наростання; калібрування по вертикалі 0,1 мВ, по горизонталі для / - 125 мс, для 2, 3, 4 250 мс. В. Імпульсна активність бистроадаптірующіхся механорецепторов (детекторів швидких швидкостей) на ступенчатообразний (/) і пилковидні (2, 3) механічні стимули з різною швидкістю наростання; калібрування по вертикалі 0,1 мВ, по горизонталі для /, 2- 250 мс, 3 75 мс зміщення шкіри (розтягнення або натискання) вони генерують потенціали дії, частота яких з часом зменшується (повільна адаптація). При збільшенні амплітуди дратівної стимулу частота аферентних потенціалів дії підвищується і залежність між силою роздратування і частотою імпульсації носить логарифмічний характер. Імпульсна активність, яка припадає на передній фронт наростання роздратування, збільшується зі збільшенням швидкості зсуву шкіри.
Паралельні морфологічні дослідження дозволили ідентифікувати деякі з цих рецепторів. Ними виявилися інкапсульовані механорецептори - тільця Руффини і тільця Меркеля, а також велика група вільних нервових закінчень різного ступеня розгалуження (рис. 15.7). Тельця Руффини розташовуються в дермі і являють собою веретеноподібні структури, оточені капсулою (див. Рис. 15.7, А). Сложнопе- реплетенние колагенові волокна є допоміжним апаратом рецептора, вони тісно пов'язані з колагеновими пучками, що лежать поруч з тільцем. Міелінізірованних волокно діаметром 5 ... 10мкм проникає всередину капсули, втрачає міелі- нову оболонку і багаторазово ділиться, утворюючи кустіковідние закінчення, розташовані на колагенових волокнах. У тілець Руффини найбільш виражена електрична реакція на розтягнення шкіри. Тельця Меркеля активніше реагують на тактильне (натиснення на шкіру) роздратування. Ці структури зустрічаються в волосистої шкірі і являють собою куполоподібні освіти діаметром 100 ... 500 мкм, що підносяться над поверхнею шкіри (див. Рис. 15.7, Б). Рецепторні закінчення локалізуються безпосередньо під базальноїмембраною утолщенного епідермісу. Міелінізірованних волокно (середній діаметр 9,5 мкм) також багаторазово ділиться і утворює дископодібні розширюються закінчення з тісно пов'язаною з ним кліткою Меркеля. Нервове закінчення містить велику кількість мітохондрій.
Крім медленноадаптірующіхся механорецепторов в шкірі були виявлені бистроадаптіруюшіеся рецептори, які, як правило, не володіють спонтанною активністю. На постійний зсув шкіри вони не генерують потенціали дії, який би величини воно не було (див. Рис. 15.6, Б). Генерація потенціалів дії відбувається тільки під час зміни амплітуди наростання або спаду стимулу. Зі збільшенням швидкості зміни амплітуди роздратування число потенціалів дії, а отже, і їх середня частота збільшуються. Ці механорецепторів були класифіковані як детектори швидкості. Бистроадаптіруюшіеся відповіді були зареєстровані від нервових волокон, що іннервують волосяні цибулини різних типів волосся: пухових, стрижневих і вибрисс. нервові оконча-
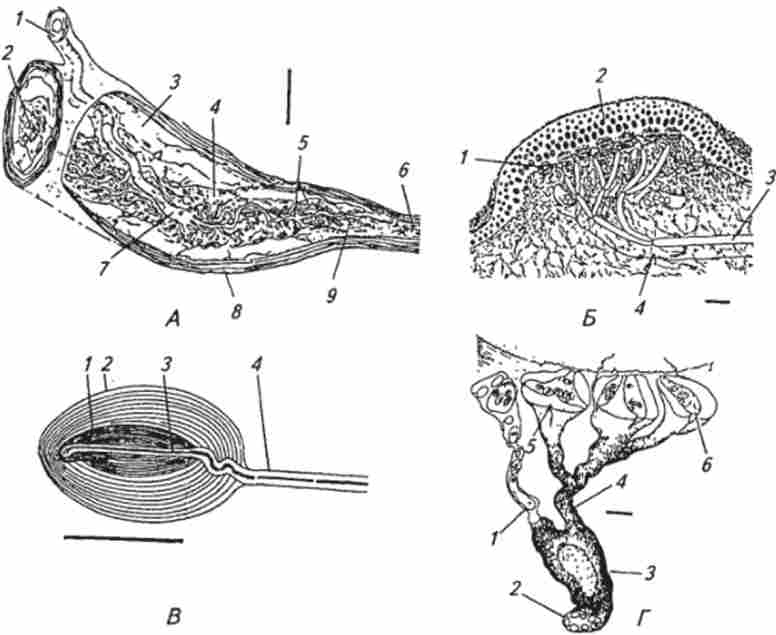
Мал. 15.7. Шкірні механорецептори (графічна реконструкція по серійним зрізах):
А. Шкіряний рецептор розтягування - тільце Руффини: / - афферентное нервове волокно; 2, 5 аферентні нервові закінчення; 3 - внутрикапсулярная простір; 4, б-колагенові волокна; 7-місце розгалуження афферентного нервового волокна; 8- капсула; 9- шванновскими клітина з ядром; калібрування 50 мкм. Б. Тактильний тільце - тільце Меркеля: / - тактильна клітина (клітина Меркеля) і примикає до неї дископодібні нервове закінчення; 2 - епідерміс тактильного тільця; 3 - афферентное нервове волокно; 4 неміелі- нізірованних нервове волокно; калібрування 10мкм. В. Тельці Пачіно: / -внутрішнє частина капсули з найбільш щільним розташуванням сплощених шарів клітин (внутрішня колба); 2 - зовнішня частина капсули з менш щільним розташуванням шарів сплощених епітеліальних клітин; 3 - афферентное нервове закінчення; 4 - афферентное нервове волокно; калібрування 600 мкм. Г. Вільні неміелінізірованние нервові закінчення (7 закінчень), утворені розгалуженням міелінізірованних нервового волокна і розташовані близько пухової волосся шкіри: / - одне з аферентних нервових закінчень (поздовжній зріз); 2-сім нервових закінчень, покритих шванівської кліткою (поперечний зріз); 3 - шванновскими клітина; 4 чотири нервових закінчення, покриті шванівської кліткою; 5, б-термінальні частини нервових закінчень, оточені складками з шванновских клітин (поперечний зріз): нервові закінчення заповнені дрібними мітохондріями; калібрування 1 мкм ня, що знаходяться в волосяних фолікулах пухових і стрижневих волосся, являють собою мережу тонких неміелінізіро- ванних закінчень, щільно обплітають структури волосяної сумки. Рецептори власний сумки утворені розгалуженими (див. Рис. 15.7, Г) міелінізірованние нервовими волокнами діаметром близько 6 мкм. Кожна волосина інервується відгалуженнями декількох волокон. У свою чергу, один і той же волокно може іннервувати багато волосся. Так, наприклад, в вусі кролика одне волокно може давати аферентні гілочки приблизно до 300 волосяних фолікулів на площі близько 1 см2. Вібриси є спеціальними дотикальними волоссям і зазвичай довший і товщі, ніж інші волосся. У коней і корів вібриси - це довгі стирчать волосся на морді; у кішок, собак і хутрових звірів їх називають вусами. У лазающих тварин вони знаходяться на животі. Корінь вібриси оточений кровоносних синусом. Вібриси сильніше іннервіровани, ніж звичайні волосся. Понад 100 нервових волокон можуть проникати в сумку вібриси і утворювати всередині аферентні закінчення. Крім бистроадаптіру- ющихся механорецепторов у вибрисс є і медленноадапті- рующим механорецептори. Детектори швидкості були виявлені в різних ділянках шкіри тварин, в тому числі в шкірі репродуктивних органів самців і сосків молочних залоз. Морфологічно вони представляють собою неінкапсулірованние нервові закінчення різної форми і складності розгалуження.
Яка ж природа адаптації в цих Механорецептори? Треба відзначити, що від детекторів швидкості не вдалося зареєструвати рецепторний потенціал, тому про причини адаптації не можна сказати щось певне. Деякі дослідники вважають, що адаптація обумовлена особливими механічними властивостями допоміжного апарату рецепторів, який не пропускає статичну компоненту механічного стимулу. Так йде справа з бистроадаптірующімся Механорецептори - тільцем Па- чини. Цей рецептор реагує на досить швидкі зміни швидкості зсуву шкіри всього лише 1 ... 2 потенціалами дії. Даний тип рецепторів отримав назву детекторів швидких швидкостей, або детекторів поштовхів.
Тільце Пачіно складається (див. Рис. 15.7, В) з нервового закінчення, мембрана якого містить механоактівіруемие іонні канали, і навколишнього його капсули у вигляді концентричних шарів з сплощені клітин епітелію, подібних верствам лусочок в цибулині. Тельці має вельми великі розміри і може досягати 0,5 ... 0,7 мм в діаметрі і 1 мм в довжину. Коли тільце при натисканні на нього ступенчатообразним механічним стимулом деформується, ця деформація через капсулу механічно передається до мембрани нервового закінчення. Остання зазвичай реагує короткочасної деполяризацией (рецепторний потенціал), що припадає на початок і кінець роздратування (рис. 15.8, А). Якщо зняти максимально можливу кількість оболонок (повністю видалити оболонки капсули нікому не вдавалося), то рецепторний потенціал буде наближатися до ступенчатообразной формі (див. Рис. 15.8, Б). Була запропонована механічна модель капсули, що представляє її шари як паралельно і послідовно з'єднані в'язко-еластичні елементи. Аналіз показав, що тиск статичної (постійної) частини стимулу послаблюється
Мал. 15.8. Електричні відповіді тактильного тільця Пачіно на ступенчатообразное механічне подразнення:
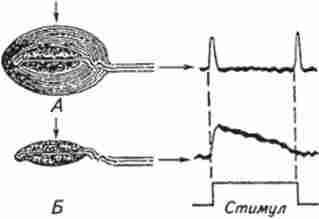
А. Интактное тільце Пачіно. На початок і кінець роздратування генерується короткочасний деполяризационного рецепторний потенціал Б. У тільця Пачіно механічно видалені зовнішні шари капсули. На механічний стимул генерується більш тривалий рецепторний потенціал
приблизно в 100 разів при досягненні області аферентного закінчення. Тільки швидкий компонент, т. Е. Зміна швидкості роздратування, може проходити через капсулу. Таким чином, капсула грає роль механічного фільтру. Треба відзначити, що в шкірі корів, кіз, коней та інших тварин було виявлено велику кількість мов теля те Пачіно, так і інших подібних їм рецепторів з різними розмірами і кількостями шарів капсули.