| Головна |
| ЗМІСТ | »» |
|---|
В результаті вивчення даного розділу студенти повинні:
знати
вміти
володіти
- принципами організації швидкої і повільної регуляції тканинного кровотоку.
Серцево-судинна система організму вирішує кілька дуже важливих взаємопов'язаних завдань. По-перше, вона забезпечує всі органи і тканини кров'ю в кількості, що точно відповідає їх мінливих метаболічним потребам. По-друге, вона здійснює перерозподіл крові між органами таким чином, щоб підвищений кровообіг активно працюють органів було скомпенсировано за рахунок працюючих менше активно і не виникало надмірних навантажень на серце. По-третє, вона підтримує при будь-яких фізіологічних станах організму такий градієнт тиску, який необхідний для нормального кровотоку і при цьому безпечний для серця і судин. Якщо перше завдання може бути вирішена за допомогою регуляторних факторів, вироблених самої споживає кров тканиною {Місцева регуляція кровотоку), То для вирішення другої і третьої задач необхідні узгоджені зміни периферичного опору великих судинних регіонів і роботи серця. Такі узгоджені реакції може забезпечити тільки НС за підтримки ендокринної. Ці механізми відносять до системної регуляції кровообігу. Залежно від органу і розв'язуваної регуляторної завдання місцеві і системні механізми можуть доповнювати один одного або надавати протилежну дію на судини.
Мішенню всіх регуляторних впливів є гладка мускулатура в складі стінок судин.
Гладкі м'язи судин складені з одноядерних дрібних клітин (міоцитів), що мають форму веретена. У великих кровоносних судинах гладеньких м'язів утворюють кругові або спіральні шари, а в артерио- лах - один круговий шар. Ендотеліальні клітини дають в гладком'язових шар численні вирости, за допомогою яких здійснюється взаємодія ендотеліоцитів і міоцитів (рис. 8.1).
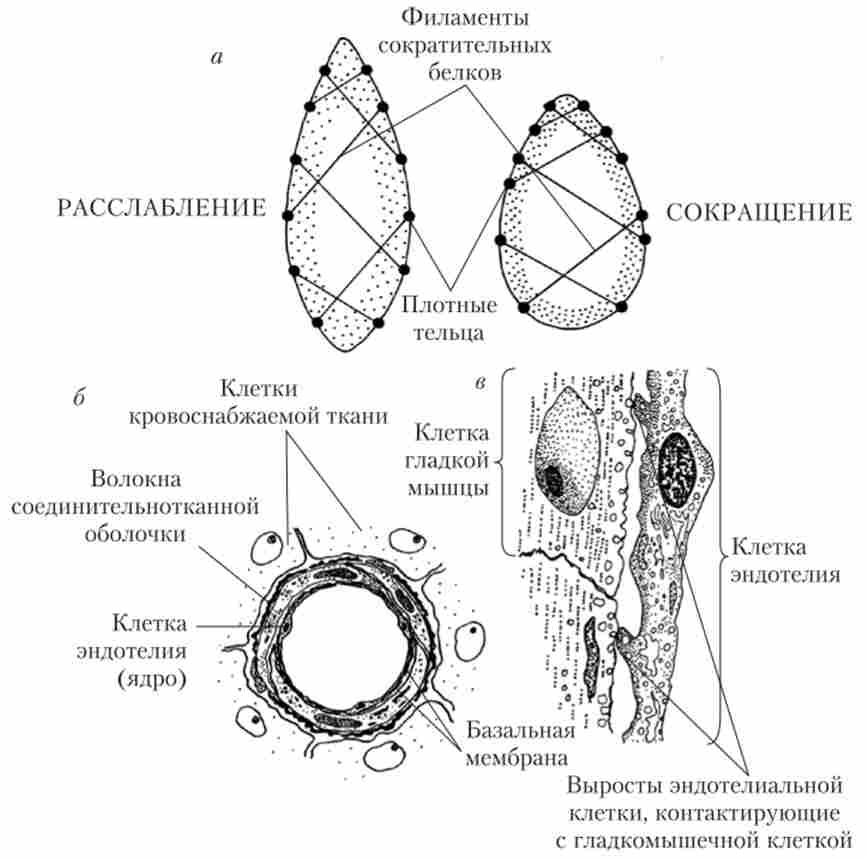
Мал. 8.1. Особливості будови гладком'язових клітин стінки судини:
а - схема закріплення на внутрішній поверхні мембрани гладком'язових клітин филаментов скорочувальних білків; б - поперечний зріз артеріоли (малюнок з мікрофотографії); в - контакти між ендотеліальними клітинами
і клітинами гладкої мускулатури
Гладеньких м'язів стінок судин мають багато структурних та функціональних відмінностей як від кардіоміоцитів, так і від поперечносмугастих волокон скелетної мускулатури. Зовнішня мембрана гладких клітин не утворює каналів Г-системи, що можна пояснити як малими розмірами міоцитів, так і тим, що ПД не є єдиними або головними сигналами для їх скорочення. Скоротливі білки формують в цитоплазмі гладких клітин велика кількість тонких (Актинові) і порівняно невелика кількість товстих (миозинових) філаментів (ниток). Видимих саркомерів з поперечними смужками ці філаменти не утворюють. Вони прикріплюються до внутрішньої сторони зовнішньої мембрани і витягуються уздовж довгої осі клітини. Скорочення гладких клітин, як і клітин інших м'язів, забезпечується ковзанням ниток актину уздовж ниток міозину, що йде з витрачанням АТФ. Однак, у порівнянні зі скелетної м'язом, гладка м'яз скорочується повільніше, розвиває велику силу і здатна залишатися в скороченому стані тривалий час при дуже незначному споживанні АТФ.
Як і в клітинах скелетних м'язів, процес скорочення в клітинах гладеньких м'язів запускають іони Са2+, проте вони взаємодіють ні з тро- поміозіном (його в гладких м'язах немає), а з білком кальмодулином, який, в свою чергу, утворює комплекс з ферментом протеїнкіназою і активує головки міозину, ініціюючи утворення поперечних містків з актином. Підвищена внутрішньоклітинна концентрація Са2+, необхідна для скорочення, виникає як за рахунок надходження Са2+ із зовнішнього середовища, так і за рахунок його викиду з саркоплазматичного ретикулума. З зовнішнього середовища він надходить через потенціалзалежні Са2+-канали (в меншій мірі) і через рецеітор-керовані Са2+-канали (в більшій мірі). З саркоплазматичного ретикулуму Са2+ викидається під впливом речовини інозитол-трифосфату, який утворюється з фосфоліпідів мембрани під впливом ферменту фосфоліпази С. фосфоліпаза З активується мембранним G-білком, пов'язаних з хімічними рецепторами. Ці ж рецептори керують мембранними Са2+-каналами. Зниження концентрації внутрішньоклітинного Са2+ при розслабленні відбувається за рахунок його активної закачування в саркоплазматичний ретикулум (Са2+-насос) і виведення з клітини. З клітини в зовнішнє середовище Са2+ виводиться за допомогою Са2+-насоса і Ка+/ Са2+-обмінника. Описані механізми представлені схематично на рис. 8.2.
Гладеньких м'язів в складі стінок судин зазвичай знаходяться в частково скороченому стані, а самі судини - частково звуженими. Це явище називається судинним тонусом.
Тонус судин виникає під впливом постійно надходять по симпатичних нервових волокнах судинозвужувальних сигналів (нейрогенний тонус), а також під впливом постійно циркулюють в крові судинозвужувальних агентів (гуморальний тонус). Крім цього, клітини гладеньких м'язів здатні скорочуватися під впливом різниці тисків по обидва боки стінки судини. Цей компонент тонусу називається базальним тонусом. У різних органах базальний тонус судин може сильно відрізнятися: він досить високий в судинах скелетних м'язів і дуже низький в шкірних судинах (у них тонус нейрогенной природи). Для артеріол, метартеріол і прекапілярнихсфінктерів багатьох органів характерні ритмічні коливання базального тонусу, що викликають ритмічні коливання швидкості кровотоку. В основі цих реакцій лежить здатність гладком'язових клітин до спонтанних ритмічним скороченням (автоматии). Механізм автоматии схожий з таким у клітинах серцевого м'яза, а його можливе фізіологічне значення полягає в постійній зміні «працюють» ділянок капілярної мережі.
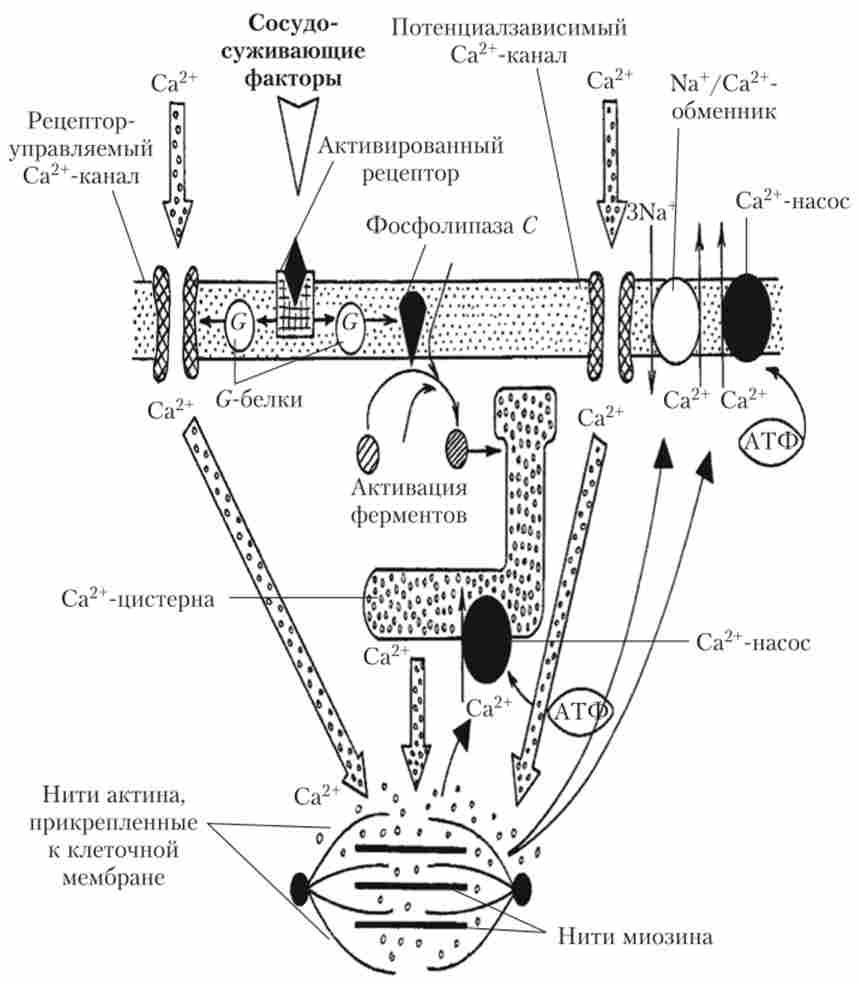
Мал. 8.2. Схема механізму процесів, що призводять до скорочення і розслаблення гладком'язових клітини:
товстими стрілками показані шляхи надходження іонів Са2+ до міофіламенти, тонкими - шляхи їх видалення з цитоплазми (подробиці див. в тексті)