| Головна |
| «« | ЗМІСТ | »» |
|---|
Поперечно-смугаста м'язова тканина утворює м'язи голови, тулуба, кінцівок, глотки, гортані, жувальних м'язів, мови, краниального відділу стравоходу. За рахунок скелетної м'язової тканини тварина може здійснювати довільні руху.
Кістякова м'язова тканина розвивається з міотомів сегментированного відділу мезодерми, покреслена м'язова тканина внутрішніх органів - з спланхнотома. Розрізняють чотири стадії формування м'язового волокна: міобла- стічний, міосімпластічну, м'язових трубочок, дефинитивного м'язового волокна.
На ранній стадії розвитку міотоми складаються з щільно розташованих епітеліоподобних м'язових клітин, які диференціюються в проміобласти, а потім в міобласти - міобласті- чна стадія розвитку (рис. 39). Ядра міобластів великі, містять хроматин і ядерця. Клітини активно діляться і переміщаються як єдине ціле потоками в ділянки розташування майбутніх м'язів. Цитоплазма міобласти спочатку має тонковолокнисту будова, потім з'являються поодинокі скорочувальні нитки.
Міобласти діляться без поділу цитоплазми, тому вони стають багатоядерними (міосімпластічна стадія). Багатоядерні структури набувають подовжену форму і утворюються м'язові трубочки. У центральній частині м'язових трубочок в ряд розташовуються численні ядра, на периферії диференціюються міофібрили. Після того як більша частина м'язових трубочок заповнюється миофибриллами, встановлюються нейром'язові контакти.
формування дефінітивних м'язових волокон супроводжується різким збільшенням числа міофібрил, що займають центральне положення, тоді як численні ядра переміщаються на периферію і розташовуються під плазмолеммой.
Частина міобластів, які не долучилися до утворення м'язових волокон, диференціюються в міосателліти - камбіальні клітини, які розташовуються між базальноїмембраною і сарколеммой.
Зростання м'язів в постнатальний період обумовлений не збільшенням числа, а потовщенням волокон. Міофіламенти будуються на поверхні вже наявних міофібрил. М'язові волокна ростуть в довжину за рахунок прибудови нових саркомерів, при пошкодженні регенерують за рахунок міосателлітов, здатних диференціюватися в міобласти і далі, як це присходят в ембріогенезі.
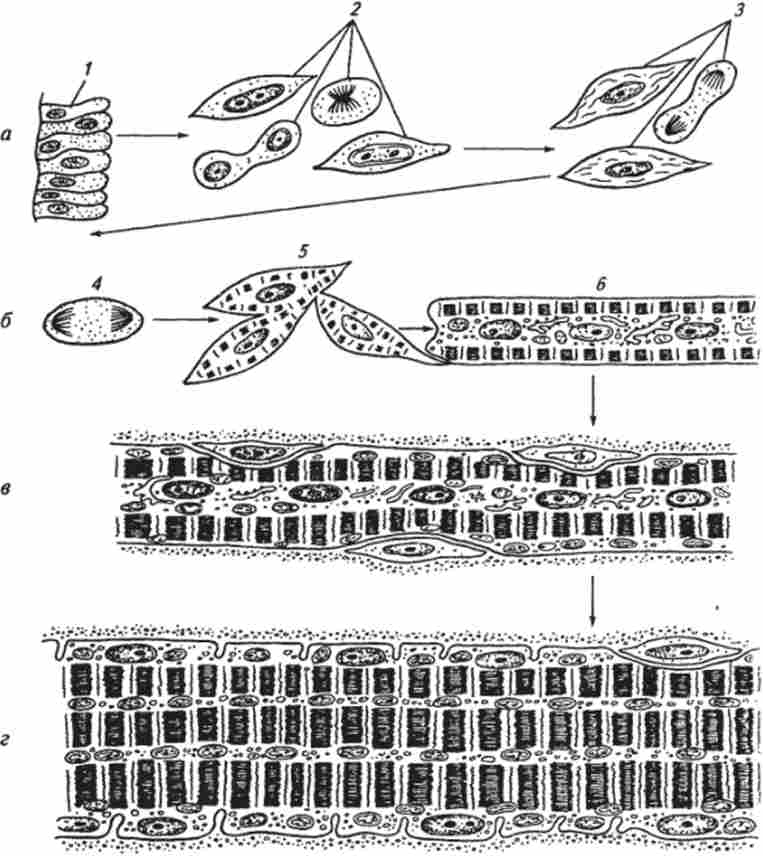
Мал. 39. Схема розвитку поперечно-смугастої скелетної м'язової тканини (по О. В. Волкової і співавт.):
а - міобластічна стадія; б - міосімпластічна стадія; в - стадія м'язових трубочок; г - стадія формування дефінітивних м'язових волокон; / - епітеліоподобная клітина міотома; 2 проміобласти; 3 міобласти; 4 - клітина в стані мітозу; 5 - злиття міо бластів і початок синтезу міофібрил; 6 - освіту миосимпластами
М'язове волокно (Міон) - структурно-функціональна одиниця скелетної м'язової тканини, має вигляд тонкого циліндра, довжиною до 13 ... 15 см, діаметром 10 ... 150 мкм. М'язові волокна можуть розташовуватися в різних напрямках: поздовжньому, косому і поперечному.
Поперечно-смугасте волокно оточує сарколеммой (від гр. Sarcos - м'ясо, lemma - оболонка). Під сарколеммой розташовуються овальні, містять хроматин ядра. Поздовжня ісчер- ченность залежить від фибриллярного будови волокна, в якому окремі фібрили тягнуться паралельно. Поперечна ис креслення визначається тим, що фібрили неоднорідні на всьому протязі: в них все строго закономірно, у всіх на одному і тому ж рівні чергуються поперемінно світлі і темні смужки (рис. 40).
Сарколеммой складається з двох шарів, утворених мембранами: зовнішня (базальна) і внутрішня (плазматична) мембрани розділені простором 10 ... 25 нм.
Базальна мембрана, розташована між пухкою волокнистою сполучною тканиною і м'язовим волокном, є посередником і місцем прикріплення ніжною мережі колагенових волокон. Елементи пухкої волокнистої сполучної тканини, розташовані зовні від базальної мембрани, утворюють ендо- мізій, навколишній окремі м'язові волокна, об'єднуючи їх в первинні пучки, що сприяє інтеграції зусиль при скороченні.
плазматична мембрана (плазмолемма) М'язового волокна безпосередньо обмежує вміст саркоплазми, виконує функцію бар'єру, що забезпечує виборчий обмін речовин між м'язовим волокном і навколишнім середовищем.
Саркоплазмою називають цитоплазму м'язового волокна. Між структурами волокна розташовано основне речовина саркоплазми - саркоплазматичний матрикс, що складається з глобулярних білків і пігментного білка - міоглобіну, здатного зв'язувати кисень.
У саркоплазме численні ядра розташовуються безпосередньо під сарколеммой, розмір і форма ядер залежать від ступеня скорочення м'язового волокна. Між миофибриллами знаходиться безліч мітохондрій - саркос.
Цитоплазма - саркоплазма м'язового волокна, містить мережу внутрішніх мембран - саркоплазматичний ретикулум. Поперек волокна між миофибриллами проходить система трубочок - Т-система, пов'язана з сарколеммой. Комплекс з однієї Т-трубочки, саркоплазматичної мережі і термінальних цистерн
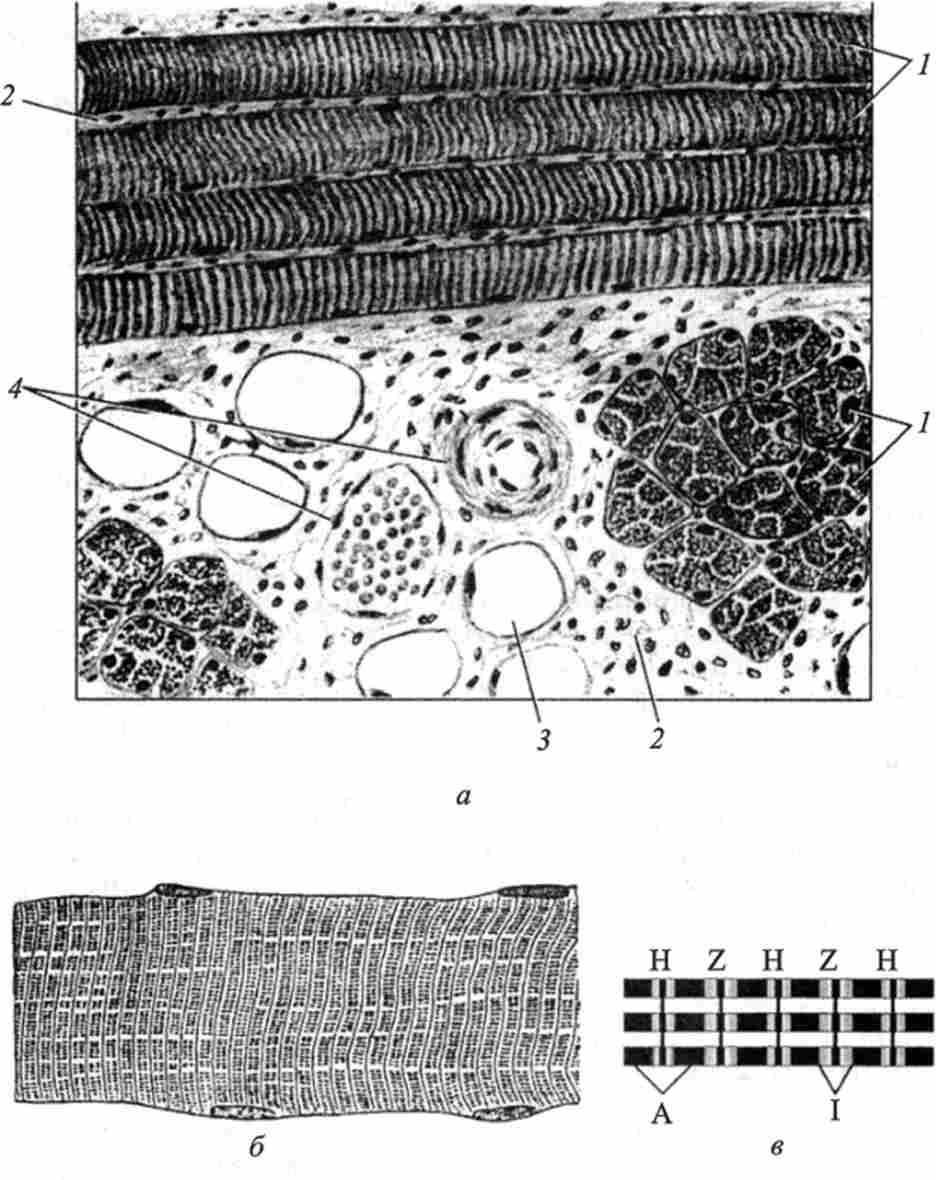
Мал. 40. Поперечно-смугаста скелетна м'язова тканина (по О. В. Волкової і співавт.):
про - поздовжній і поперечний розрізи; б- м'язове волокно; в - структура міофіб- Рилл: A-диск; 1-диск; Z-смужка; Н-смужка; / - м'язові волокна; 2-прошарки пухкої сполучної тканини; 3 - жирові клітини; 4 - кровоносні судини називають тріадою. Тріада бере участь в просуванні хвиль деполяризації, акумуляції та вивільнення іонів кальцію. В результаті концентрація цих іонів в саркоплазме знижується або підвищується, що, в свою чергу, впливає на активність АТФази, отже, на скоротливу функцію м'язового волокна.
Міофібрили - скорочувальної апарат м'язового волокна - це поздовжньо орієнтовані білкові скорочувальні нитки, що мають характерну смугастість.
Смугастість міофібрили обумовлена чергуванням темних і світлих дисків, які по-різному заломлюють поляризоване світло.
Світлі, ізотропні 1-диски мають одинарним; темні, анізотропні А-диски - подвійне променезаломлення (див. рис. 40).
Темні і світлі диски міофібрил розташовані впорядковано один навпроти іншого, що і надає волокну поперечну смугастість (рис. 41). У темних дисках знаходяться товсті міо Зіновій нитки, що містять світлу Н-смужку, в середині якої проходить темна М-лінія.
Світлі диски містять тонкі (актинові) нитки, які посередині перетинає Z-смужка. Ділянка міофібрили між сусідніми Z-смужками - саркомер.
Саркомер - структурно-функціональна одиниця міофібрили, розташована між Z-смужками; отже, кожен саркомер містить один A-диск і дві половинки 1-диска. Один кінець тонкої (актиновой) нитки прикріплений до Z-смужці, а інший направлений до середини саркомера, т. Е. Тонкі нитки проходять між товстими миофиламентами в область A-диска, з якими пов'язані поперечними містками, що відходять від товстих миофиламентов через кожні 60. ..70 нм.
На поперечних зрізах виявляють гексагональную структуру ниток: одна товста нитка оточена шістьма тонкими, відстань між нитками варіює від 20 до 30 нм. Міофібрили виявляють у вигляді точок, іноді заповнюють волокно рівномірно, іноді вони розділені прошарками саркоплазми, утворюючи міо фібрилярні поля Конгейма, кожне з яких відповідає поперечному розрізу міофібрил.
Довжина товстих ниток, відповідно, довжина A-диска дорівнює 1,6 мкм; тонкі нитки тягнуться на 1 мкм в обидві сторони від Z-смужки. Це означає, що в стані спокою довжина саркомера - 2,6 мкм, отже, область перекриття буде по 0,5 мкм в кожній з половинок саркомерів.
Актинові філаменти мають діаметр 5 нм. Вони складаються з актину, тропомиозина і тропоніну. Кожна актинового нитка утворена двома спіральними тяжами з глобулярних молекул акти-
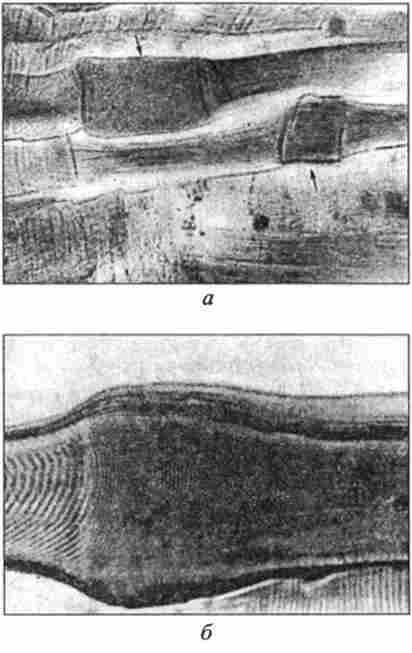
Мал. 41. Вузли скорочення м'язових волокон, збільшення х280; 400 (по Г. Г. ТІНЯКОВА):
а - кашалота; б - великої рогатої худоби
на, закручених один навколо іншого. Весь комплекс актінових молекул називається фібрилярні актином (F-актин). З кожною молекулою F-актину пов'язана молекула АТФ.
Міозіновие філаменти мають діаметр 10 нм і складаються з білка міозину. Молекула міозину складається з двох частин: довгої ділянки - «хвоста» і приєднаного до одного з кінців глобулярного ділянки, який представлений двома однаковими «головками». У тих місцях, де нитки актину і міозину перекриваються, головки можуть прикріплятися до сусідніх Актинові ниток, в результаті цього відбувається вкорочення м'яза. Енергія для скорочення вивільняється при гідролізі АТФ, так як міозіновие головки мають АТФазной активністю.
Актоміозін - комплексний білок, при утворенні молекули якого до бусинкам актину прикріплюються головки міозину. Молекулярна маса білка коливається в широких межах, так як співвідношення актину і міозину в комплексі може бути різним. Актоміозін не розчиняється у воді, розчин відрізняється високою в'язкістю, яка залежить від співвідношення актину і міозину (чим більше актину, тим вище в'язкість). Актоміозін здатний диссоциировать в присутності АТФ і іонів магнію.
Скорочення скелетного м'яза хребетних визначаються наявністю спеціалізованих допоміжних білків. Один з цих білків - тропомиозин (Жорстка молекула), що складається з двох однакових спіральних ланцюгів, по 284 амінокислоти в кожній. Тропомиозин зв'язується з Актинові філаментів по всій довжині і додає жорсткість тонким ниткам.
тропонин являє собою комплекс з трьох поліпептидів, що забезпечують здатність міофібрил швидко реагувати на підвищення концентрації іонів кальцію. На кожні сім мономерів актину доводиться тільки один тропоніновим комплекс.
Білки м'язової тканини підрозділяють на водорозчинні, солерозчинних і нерозчинні в водно-сольових розчинах фракції. Розчинні у воді білки входять в основному до складу плазми, солерозчинних утворюють міофібрили, нерозчинні в водно-сольових розчинах фракції умовно називають білками строми, до складу яких входять білки сарколеми, ядер і внутрішньоклітинні сполучнотканинні білки.