| Головна |
| «« | ЗМІСТ | »» |
|---|
У морфологічному відношенні м'ясо являє собою складний тканинний комплекс, до складу якого входять м'язова, сполучна (пухка волокниста, жирова, кісткова) тканини, кровоносні і лімфатичні судини, нервові волокна і нервові закінчення. Залежно від типу годівлі, віку, породи та умов утримання тварини одні й ті ж м'язи можуть мати різні розміри, форму, що враховується при товарознавчої оцінки якості туш.
Важливою ознакою, що дозволяє диференціювати м'язи хребетних тварин, є розташування ядер в м'язовому волокні. У амфібій, риб, птахів ядра розташовані в центральній частині, тоді як у ссавців - по периферії волокна (рис. 42, 43, 44).
Скелетний м'яз зовні оточена оболонкою з пухкої волокнистої сполучної тканини, яка називається епімізіем. Від епімізія всередину відходять прошарки пухкої волокнистої сполучної тканини - перілшзіи, - навколишні м'язові пуч-
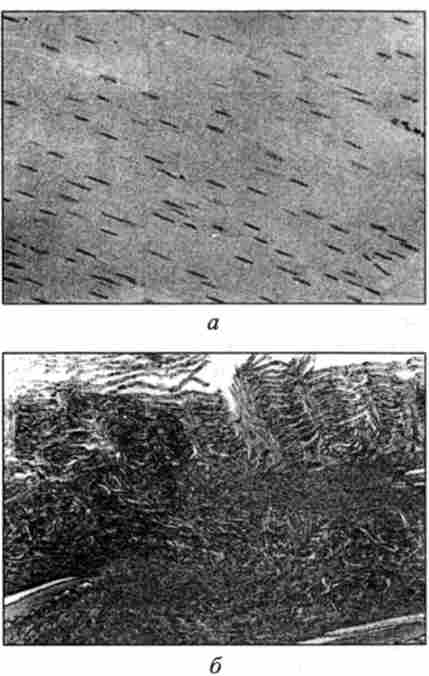
Мал. 42. Мікроструктура м'язової тканини. Забарвлення гематоксиліном і еозином,
окуляр х7; об'єктив х40:
а - жаби; 6 - коропа
ки різної величини. Між м'язовими волокнами проходять дуже ніжні і тонкі сполучнотканинні прошарки - ен домізій. Перимизий і епімізій складаються з колагенових і еластичних волокон різної міцності, що утворюють більш-менш складні сплетіння. У перимизия і епімізіі м'язів відгодованих тварин знаходяться жирові клітини, що зумовлюють мраморност' м'яса.
У сільськогосподарських тварин скелетна м'язова тканина представлена різними типами м'язових волокон, що обумовлено функціями: неоднаковою силою, швидкістю, тривалістю скорочення, і в зв'язку з цим розходженням в будові. На світлооптичному рівні розрізняють червоні, білі, проміжні, спеціалізовані м'язові волокна.
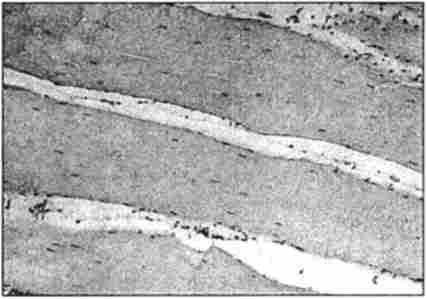
Мал. 43. Мікроструктура м'яса індички. Забарвлення гематоксиліном і еозином,
окуляр х7; об'єктив х40
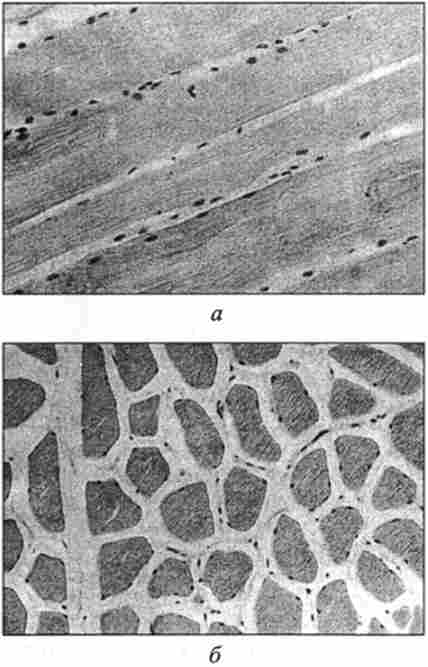
Мал. 44. Мікроструктура свинини. Забарвлення гематоксиліном і еозином, окуляр х7;
об'єктив х40:
а - поздовжній розріз; б - поперечний розріз
Червоні м'язові волокна характеризуються пухким розташуванням міофібрил, великою кількістю саркоплазми, добре розвиненим мітохондріальних апаратом і великою насиченістю ліпідами. Для червоних волокон на відміну від білих характерні повільне скорочення і висока тривалість цього процесу.
Білі м'язові волокна характеризуються щільним розташуванням міофібрил, відносно невеликою кількістю саркоплазми, значною кількістю глікогену. Ці волокна швидко скорочуються, але й швидко втомлюються.
Проміжні м'язові волокна відрізняються від червоних і білих, мають незначну скорочувальної здатністю.
У спеціалізованих м'язових волокнах (нервово-м'язове веретено) ядра сконцентровані у вигляді ланцюжка тільки по центру волокна. На таких м'язових волокнах, що мають вигляд веретена, нервові волокна утворюють аферентні закінчення. Скорочення волокон веретен не підсумовується зі скороченнями робочих м'язових волокон, вони лише натягують капсулу, забезпечуючи регуляцію м'язової діяльності.
Кількість і співвідношення м'язових волокон різних типів у тварин може змінюватися з віком. Так, у свиней в довгій м'язі спини з віком кількість білих волокон зростає, кількість проміжних волокон знижується. У червоних м'язових волокнах АТФ утворюється окислювальним шляхом, в білих - гликолизом; перші переважають в м'язах динамічного типу, а другі - в м'язах статичного і статодинамичні типів.
Після забою тварини, з припиненням обміну речовин, основним біохімічним процесом при переробці сировини є автолиз (від гр. Autos - сам, lysis - розчинення), основу якого складають ферментативні процеси, при житті пов'язані з функцією координованого руху.
Автолиз - процес розпаду речовин і тканин під дією ферментів самих тканин. Спочатку переважають гліколітичні процеси, що призводять до накопичення молочної та ортофосфор- ної кислот. Величина pH м'яса, що визначається кількістю молочної кислоти, обумовлює технологічні, мікробіологічні і товарні якості м'яса і м'ясопродуктів. У міру зміни pH створюються умови, що сприяють дії різних тканинних протеолітичних ферментів, що призводять до поліпшення консистенції м'яса.
Сукупність змін найважливіших властивостей, обумовлених розвитком автолізу, внаслідок яких м'ясо набуває ніжну консистенцію і соковитість, добре виражені специфічні запах і смак, - дозрівання м'яса.
Автолиз розвивається в певній послідовності: пос леубойное розслаблення м'язів, розвиток посмертного задубіння, дозвіл посмертного задубіння і власне дозрівання.
парне м'ясо - вихідна структура, з якою можна порівнювати всі наступні зміни в м'ясі. М'язові волокна парного м'яса мають гофровану і складчасту форми, що обумовлено різним ступенем скорочення: одні слабо скорочені або розслаблені, а розташовані поруч з ними прямі волокна сильно скорочені. Для порівняння: в охолодженому м'ясі складчастість м'язових волокон майже повністю зникає.
Ступінь скорочення м'язових волокон можна визначати за наявністю вузлів скорочення, що представляють характерну структуру, яка виникає в поперечно-смугастих м'язових волокнах всіх видів сільськогосподарських тварин.
За будовою вузли скорочення характеризуються сильним ступенем скорочення міофібрил, ступінь скорочення яких по довжині вузла часто розподіляється закономірно: найвища ступінь скорочення розташована ближче до центру вузла. У центральній частині вузлів спостерігають щільне зближення саркомерів і виникає суцільна бесструктурная темна маса. Ця маса в центрі вузлів зазвичай стає місцем роз'єднання волокна на окремі фрагменти, кількість яких збільшується, якщо вузли скорочення довгі. Вузли скорочення формуються на свежеобре- занних м'язових волокнах у вигляді «тюльпанів» (див. Рис. 41).
Характерною рисою в будові вузлів скорочення є наявність «віяла», в якому центральна частина є вузлом скорочення, периферична складається з розслаблених міофібрил, що мають гофровану структуру. Частота вузлів скорочення стає особливо високою в період посмертного задубіння (rigor mortis). Дозвіл посмертного задубіння відбувається асинхронно і виражається різним ступенем розтягнутості окремих м'язових волокон і їх міофібрил.
В окремих волокнах виявляють поодинокі або множинні вузли і смуги скорочення, що представляють собою локальні сверхсокращенние ділянки міофібрил. Утворення таких ділянок пов'язано зі змінами актінових протофібрілл в області A-дисків, що виражаються зиґзаґоподібної складчатостью.
В результаті автолізу скорочені волокна кілька розслабляються і випрямляються. Діаметр поверхнево розташованих волокон зменшується внаслідок більшої втрати води в порівнянні з волокнами, що лежать на глибині. Ядра в підсихаючих поверхневих волокнах відрізняються від ядер глубоколежа- щих волокон більшої ущільнення.
PSE-м'ясо має низьке значення величини pH (нижче 5,6) і характеризується як бліде, м'яке, водянисте. DFD-м'ясо має високе значення pH (вище 6,2) і характеризується як темне, клейка, щільної консистенції. Ступінь деформації м'язових волокон PSE- м'яса значно менше в порівнянні з цієї ж м'язом з нормальним значенням pH. У ряді волокон відзначають множинні гомогенні, безструктурні, що не мають певної форми смуги, що йдуть поперек волокон. Деструктивні зміни м'язових волокон (порушення цілісності) виявляють у вигляді множинних поперечно-щілиновидних мікротріщин.
Для DFD-м'яса ступінь деформації м'язових волокон незначна. М'язові волокна переважно прямі, з локальної деформацією окремих ділянок, щільно прилеглих один до одного; поперечна і поздовжня смугастість м'язових волокон слабо виражені. У деформованих ділянках волокон зі слабо вираженою поперечною смугастість відзначають дезорієнтацію і часткову деструкцію міофібрил. М'язова тканина DFD-м'яса має нерівномірну, плямисте забарвлення волокон.
В охолодженому м'ясі з'являються поперечні тріщини, які в парному м'ясі майже не зустрічаються. Тріщини виявляють в сильно скорочених волокнах. Основна маса таких тріщин, мабуть, виникає в період посмертного задубіння, коли більшість м'язових волокон знаходиться в сильно скороченому стані. Різна ступінь синхронності скорочення волокон призводить до виникнення в них деформацій. Світлі 1-диски поступово зменшуються в розмірі і зникають, міофібрили гомогенізуються.
При зберіганні охолодженого м'яса велике значення має освіту «корочки підсихання» в поверхневих шарах туші і висівок в зв'язку з уповільненням і припиненням розвитку мікрофлори і проникненням її в товщу м'яса. Вже через одну добу зберігання м'язові волокна в охолоджених напівтушах в значній мірі втрачають гофровану структуру.
У поверхневих волокнах поперечнасмугастість виражена значно слабкіше, в деяких місцях може не виявлятися, в волокнах глибоких шарів вона чітко виражена. Поверхневі шари м'язової тканини в міру збільшення терміну зберігання обезвоживаются і ущільнюються. Це, в свою чергу, сприяє утворенню чітко вираженою «корочки підсихання», яка охороняє шари охолодженого м'яса, що лежать в глибині, від висихання і псування.
Зміни, що виникають при заморожуванні, характеризуються появою нового структурного компонента - водних кристалів і зміною загального вигляду і товщини м'язових волокон. Кристали в м'ясі утворюються за рахунок перенесення кристалізується рідини з тканинного соку. Чим глибше розташований шар волокон, тим нерівномірніше розподіл вимерзлими води в м'ясі.
Процеси кристалізації є причиною пухкого і хаотичного розташування м'язових волокон, їх фрагментації і деформації. У глибоких шарах м'яса, замороженого при температурі -20 ° С, в зоні локалізації кристалів льоду спостерігають значні зміни: деформацію і розриви міофібрил м'язових волокон, деструкцію саркоплазматичного ретикулуму, локальний розпад сарколеми, розпушення і часткове руйнування волокнистих структур сполучної тканини. В охолодженому м'ясі утворюються більші кристали, тому більш виражено стиснення міофібрил м'язових волокон. У такому м'ясі, на відміну від парного, при утворенні кристалів льоду відбуваються більш значні відшарування і локальні деструкції сарколеми з виходом дрібнозернистої білкової маси в міжм'язової простір.
Бактеріальна контамінація (псування) починається на поверхні, а потім проникає в толше м'яса, причому швидкість розвитку процесу залежить від температури і вологості навколишнього середовища, стану «корочки підсихання» і виду бактерій (рис. 45, 46).
Мікроорганізми, що розвиваються в кислому середовищі, зрушують pH в лужну сторону, готують умови для жізнедеятель-
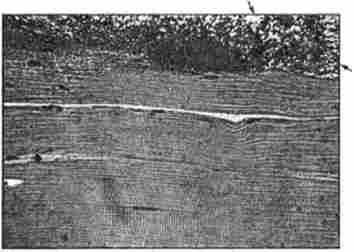
Мал. 45. Мікроструктура яловичини: на поверхні м'язових волокон виявлена мікрофлора. Забарвлення гематоксиліном і еозином, окуляр х7; об'єктив х40

Мал. 46. Мікроструктура яловичини, експериментально контамінованої бактеріями Yersinia enterocolitica. Забарвлення гематоксиліном і еозином, окуляр х7;
об'єктив х40:
а - контроль; б- досвід: в сполучної тканини між м'язовими волокнами виявлена дифузно розташована мікрофлора
ності інших мікроорганізмів. В результаті розвитку мікрофлори білок розпадається з утворенням як первинних, так і вторинних продуктів гідролізу, істотно впливають на органолептичні показники і харчову цінність м'яса.
Вивчення мікроструктури дозволяє встановити ступінь взаємозв'язку, розподіл, кількісне співвідношення і розміри складових частин фаршу. У готовому фарші виявляють: мелкозернистую масу аморфного світлішого речовини; окремі фрагменти м'язових волокон; краплі жирової речовини із зруйнованих жирових клітин, а при використанні порошкоподібних прянощів виявляють окремі фрагменти або групи клітин. У аморфному речовині зустрічаються досить великі вакуолі (порожнечі овальної і круглої форм) - це ділянки, де були повітряні бульбашки, що виникають в процесі технологічної обробки.
Підміну м'яса забійних тварин м'ясом інших хребетних тварин виявляють по положенню ядер в м'язовому волокні: у амфібій, риб, птахів ядра розташовані в центральній частині, тоді як у ссавців - по периферії волокна (див. Рис. 42, 43, 44).
У готовому продукті необхідно враховувати зміни, які відбуваються в процесі подрібнення, засолу, варіння, що відбивається на ступеня деструкції м'язових волокон, розподілі зернистої маси, стані взаємозв'язку волокнистого матеріалу, розмір і форму вакуолей і розподілі жирової фракції, наявності мікрофлори (рис. 47, 48).
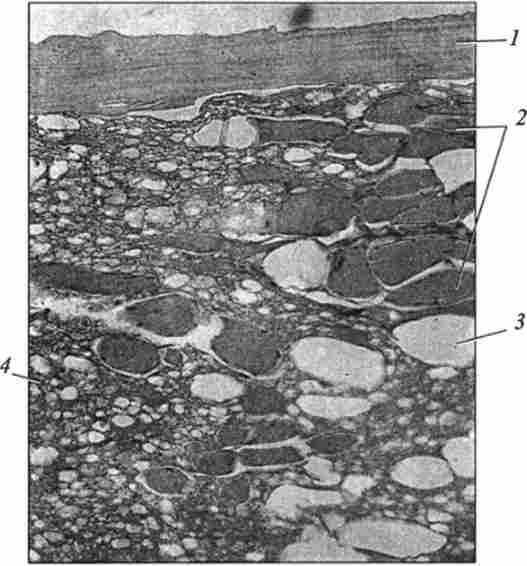
Мал. 47. Мікроструктура вареної ковбаси. Забарвлення гематоксиліном і еозином,
окуляр х7; об'єктив х40:
1 - оболонка; 2 - фрагменти м'язових волокон; 3 - вакуолі; 4 - жирові клітини
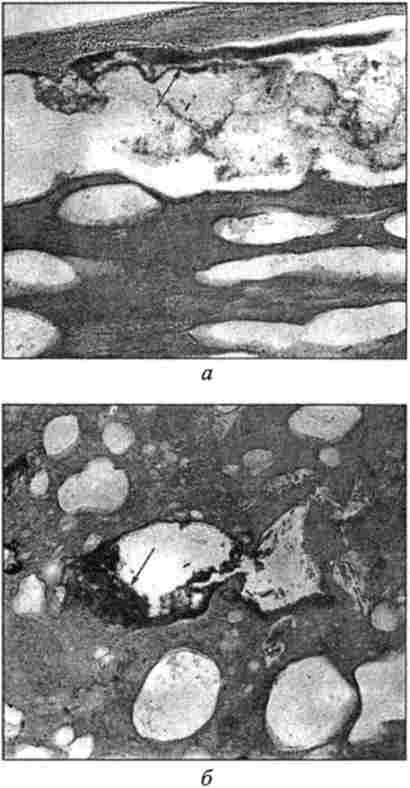
Мал. 48. Мікроструктура вареної ковбаси, експериментально контамінована бактеріями Yersinia enterocolitica. Забарвлення гематоксиліном і еозином, окуляр х7;
об'єктив х40:
а - мікрофлора під оболонкою; б- мікрофлора в вакуолях
При виготовленні варених ковбас (див. Кол. Вкл., Рис. XI) використовують м'ясну обріз, м'ясо голів, губи, вуха та ін. На гістологічних препаратах у таких випадках виявляють слинні залози, фрагменти хрящів, багатошаровий плоский епітелій слизової оболонки ротової порожнини. М'ясні консерви також можуть містити значну кількість сполучної тканини, добавки рослинних білків і крохмалю; м'язову тканину замінюють субпродуктами.
В ковбасах виявляють заміну м'яса на рослинні добавки, частіше соєві, і істотне зміст субпродуктів. Використання м'яса механічної дообвалки встановлюють по великій кількості кісткових і хрящових частинок. При використанні сировини з низькими технологічними властивостями виявляють різні білкові компоненти, наприклад соєвий ізолят, у вигляді щільних і компактних еозинофільних утворень в порівнянні з дрібнозернистої білкової масою округлої форми. Структурований соєвий білок має вигляд великих фрагментів з щільної волокнистої структурою, забарвлюється кислими барвниками. Фрагменти текстурованого білка відрізняються від м'язових волокон однорідністю та щільністю, а також відсутністю смугастість. Клітини рослинної тканини великі, чітко окреслені, мають целюлозний оболонку і слабо сприймають гістологічні барвники.
Дослідження мікроструктури дозволяє в найкоротші терміни достовірно визначити в м'ясних продуктах наявність сполучної, хрящової, кісткової тканин, субпродуктів, а також рослинні добавки.