| Головна |
| «« | ЗМІСТ | »» |
|---|
Молекули нуклеїнових кислот всіх типів живих організмів - це довгі не- розгалужені полімери мононуклеотидів. Роль містка між нуклеотидами виконує 3 ', 5'-фосфодіефірная зв'язок, що з'єднує 5'-фосфат одного нуклеотиду і З'-гід Роксі залишок рибози (або дезоксирибози) наступного. У зв'язку з цим підлогу і нуклеотидная ланцюг виявляється полярної. На одному її кінці залишається вільною 5/-0-Фн-група, на іншому З'-ОН-група.
Для запису структури нуклеїнової кислоти або її фрагмента широко використовують скорочену символіку (рис. 14.1).
ДНК, подібно білкам, має первинну, вторинну і третинну структури.
Первинна структура ДНК. Дана структура визначає закодовану в ній інформацію, представляючи собою послідовність чергування дезоксірібонукле- відійшов в підлогу і нуклеотидної ланцюга. Хоча ДНК містить всього чотири типи мономерних ланок, кількість можливих нуклеотиднихпослідовностей перевершує таке для білків внаслідок істотно ббльшсй довжини полінуклеотидних ланцюгів.
Визначення первинної структури ДНК довгі роки залишалося нерозв'язним завданням. У 70-х рр. XX ст. з відкриттям ферментів рестриктаз, «ріжуть» молекули ДНК в суворо визначених точках, А. Максамом і В. Гилбертом був розроблений метод секвенування, що дозволяє визначати послідовності до 1000 нуклеотидів. У 1985 р вдалося створити прилад для автоматичного аналізу нуклеїнових кислот. В останнє десятиліття в цій галузі намітився суттєвий прогрес, в результаті чого визначено послідовність не тільки окремих генів, але і цілих хромосом у різних видів живих організмів, в тому числі і людини. Всі дані по структурі генів, що публікуються в світовій науковій літературі, вводяться в пам'ять комп'ютера, формуючи банк даних.
В результаті проведених досліджень було встановлено, що в молекулах ДНК бактеріофагів майже всі послідовності нуклеотидів унікальні, т. Е. Зустрічаються один раз. У ДНК бактерій більшість генів також унікальні, але деякі послідовності (кодують транспортні та Хвороби) повторюються по декілька разів. У геномі еукаріотів унікальні послідовності нуклеотидів, т. З. структурні гени, що несуть інформацію про структуру специфічних білків, складають близько 60% ДНК. Іншу частину ДНК складають повторювані послідовності. Від 10 до 25% генома тварин представлено помірно повторюваними послідовностями. Вони є структурними генами продуктів, необхідних клітці в великих кількостях. Це гени рибосомних і транспортних РНК, білків гістонів, окремих ланцюгів імуноглобулінів. Вони, як правило, розташовані в ДНК у вигляді тандемних повторів, т. Е. Один за одним, один ген відділяється від іншого спейсером (від англ, spacer- проміжок). До групи помірно повторюваних послідовностей входять також ділянки ДНК, що виконують регуляторні функції. Крім того, в ДНК еукаріот зустрічаються часто повторювані послідовності (105- 106раз). В основному це сате- літної ДНК, що виявляється в центромерних областях хромосом, що бере участь, мабуть, в спарюванні і розбіжності хромосом.
Незважаючи на відмінності в первинній структурі ДНК, в сумарному нуклеотидном складі всіх типів ДНК є загальні закономірності, встановлені Е. Чаргафф, які підтверджені величезним фактичним матеріалом, який зіграв важливу роль у формуванні уявлення про вторинної структурі ДНК.
Закономірності Чаргаффа зводяться до наступного:
У ДНК деяких видів переважає сумарну кількість аденіну і тиміну, це так звані АТ-тип ДНК. АТ-тип переважає у всіх хребетних і безхребетних тварин і вищих рослин. ГЦ-тип (з сумарним переважанням гуаніну і цитозину) зустрічається у мікроорганізмів, хоча деякі з них можуть мати і АТ-тип. У зв'язку з цим Е. Чаргафф висунув положення про видовий специфічності ДНК по нуклеотидному складу. Нуклеотидний складу ДНК бактерій в даний час використовують як один з таксономічних ознак.
Вторинна структура ДНК. Відповідно до моделі, запропонованої в 1953 р Дж. Уотсоном і Ф. Криком, вона являє собою двухцепочечную правозакрученной спіраль з комплементарних один одному антіпараллсльних полінуклеотидних ниток.
Для вторинної структури ДНК вирішальним є дві особливості будови азотистих основ нуклеотидів. Перша полягає в наявності груп, здатних утворювати водневі зв'язки. Так, між А і Т можуть утворюватися дві, а між Г і Ц - три водневі зв'язку. Ці азотисті основи називаються комплементарними. Друга особливість полягає в тому, що пари комплементарних основ А-Т і Г-Ц виявляються однаковими не тільки за розміром, але і за формою (рис .14.2).
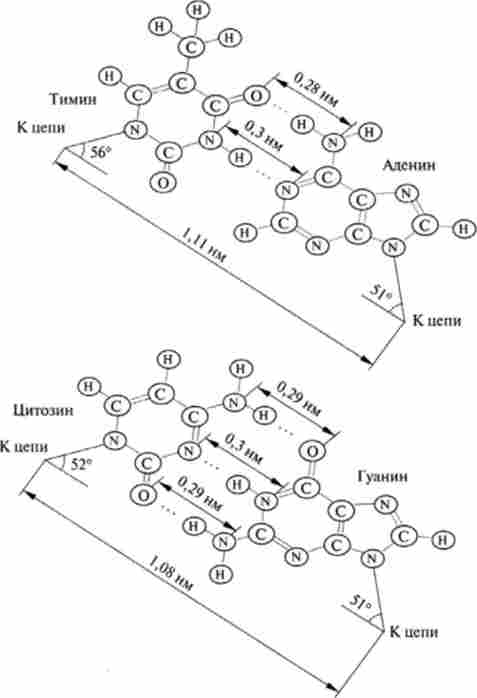
Мал. 14.2. Пари підстав, пов'язані водневими зв'язками: для А-Т і Г-Ц-пар міжатомні відстані та кути приблизно однакові
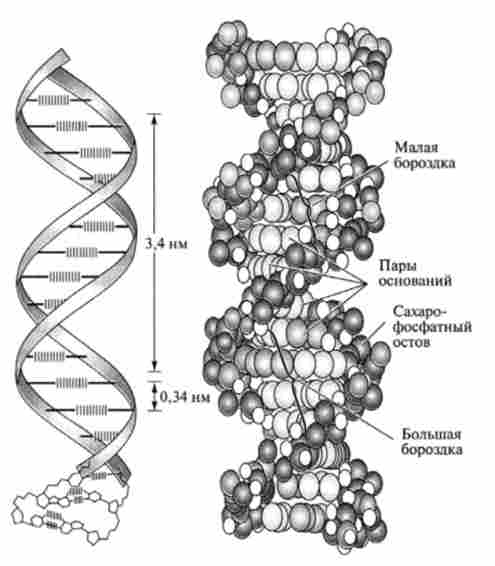
Мал. 14.3. Подвійна спіраль ДНК
Завдяки здатності нуклеотидів до спаровування, утворюється жорстка, добре стабілізована дволанцюжкова структура, що володіє наступними властивостями (рис. 14.3).
На основі ретельного аналізу рентгенограм виділених ДН До встановлено, що подвійна спіраль ДНК може існувати у вигляді кількох форм (А. В, С, Z і ін.). Зазначені форми ДНК розрізняються діаметром і кроком
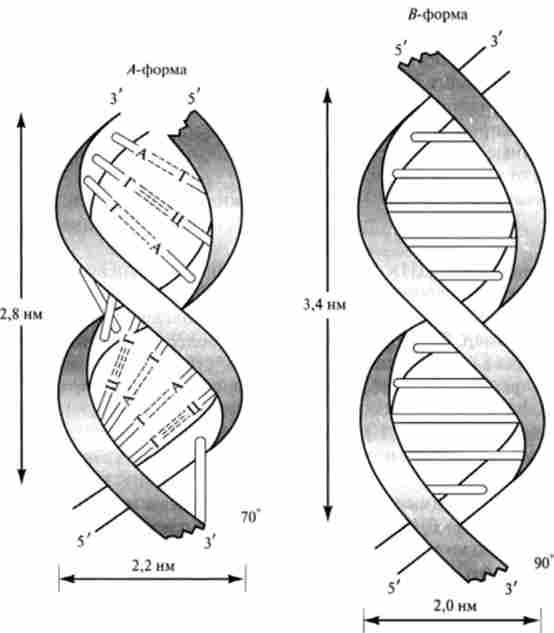
Мал. 14.4. схема А- і 5-форм подвійної спіралі
спіралі, числом пар основ в витку, кутом нахилу площини підстав по відношенню до осі молекули (рис. 14.4).
В формі В, описаної моделлю Дж. Уотсона і Ф. Кріка, на один виток спіралі доводиться 10 пар основ, крок спіралі 3,4 нм, діаметр 1,8 нм, кут нахилу до осі 0 °. форма В, мабуть, сприятлива для процесу реплікації. В формі А на один виток припадає 11 пар основ, крок спіралі 2,8 нм, кут нахилу на площині підстав до осі становить 20е. форма А є кращою для процесів транскрипції. Форма С, виявлена у ряду вірусів і в складі надмолекулярних структур хроматину, має 9,3 пари підстав в витку з кутом нахилу 5е.
Z-Форма ДНК - найменш скручена (12 пар основ на виток). Вона являє собою левозакрученной подвійну спіраль, в якій фосфо ефірний остов розташований зигзагоподібно уздовж осі. Z-Форма володіє тільки однією борозенкою. Як відомо, в борозенках ДНК регуляторні білки можуть специфічно взаємодіяти з певними атомами нуклеїнових підстав, т. Е. «Дізнаватися» конкретні нуклеотидні послідовності без порушення комплементарних взаємодій в структурі подвійної спіралі. Тим самим регуляторні білки можуть здійснювати контроль експресії генів. Деякі білки, що зв'язуються в великої чи малої борозенках форми, ймовірно, не здатні зв'язуватися з Z-формою. У зв'язку з цим Z-форма, що виникає, як правило, при високих концентраціях солей, сперміну, спермидина, при метилировании залишків дезоксіцітідіна, при високому вмісті негативних супер витків в молекулі ДНК, може брати участь в регуляції експресії генів.
Описані форми ДНК здатні до взаємно оборотних переходах в залежності від умов середовища.
Третинна структура ДНК. У всіх живих організмів двоспіральні молекули ДНК щільно упаковані з утворенням складних тривимірних структур.
Дволанцюжкові ДНК прокаріот, мають кільцеву ковалентно-замкнуту форму, утворюють ліві (-) суперспирали. Суперспіралізації перш за все необхідна для «упаковки» величезної молекули ДНК в малому обсязі клітини. Наприклад, ДНК?. coli має довжину більше 1 мм, в той час як довжина клітини не перевищує 5 мкм. Крім цього, суперспіралізації ДНК, що полегшує її розплітання, забезпечує початок реплікації і транскрипції (рис. 14.5).
Третинна структура ДНК еукаріотів також утворюється шляхом суперспіралізації, але не вільної ДНК, а її комплексів з білками хромосом.
Ядерний хроматин містить ДНК, гістонові і негістонові білки, невелика кількість РНК. В просторової організації хромосом можна виділити кілька рівнів. Перший рівень - нуклеосомної. Нуклеосом- ная нитка утворюється при взаємодії ДНК з білками-гистонами. Гістони представляють собою прості білки з молекулярною масою 14-20 kDa, в амінокислотним складом яких переважають аргінін і лізин, гліцин і цистеїн. Переважання лізину і аргініну надає гістонів лужний характер і забезпечує їх здатність взаємодіяти з кислотними групами ДНК. У всіх типах еукаріотичних клітин виявлено 5 класів гістонів
Мал. 14.5. Третинна структура ДНК прокаріотів:
а - лінійна олноцепочечная ДН К - бактеріофаг ф х 174 та інші віруси: б - кільцева одноцепочечная ДНК вірусів і мітохондрій; в - кільцева подвійна спіраль ДНК
(Н1, Н2, НЗ, Н4, Н5), рахтічаюшіхся за змістом (%) основних амінокислот, що обумовлює їх фізико-хімічні властивості (електрофоретична рухливість, ВЕТ і ін.). Гістони є еволюційно консервативними білками. Ступінь гомології амінокислотних послідовностей гістонів Н2, НЗ, Н4, Н5 у різних видів тварин, рослин і грибів досить висока. Ці гистони попарно утворюють октамер (білкові кори дисковидной форми, які обплітаються молекулою ДНК). Ділянка ДНК, спірально обплітає октаметр, містить в середньому 145-150 нуклеотидних пар і формує приблизно 1,75 витка лівої спіралі. Вільні від контакту з білковими корами ділянки ДНК називають лінкерних (або сполучними). Їх довжина варіює в за-
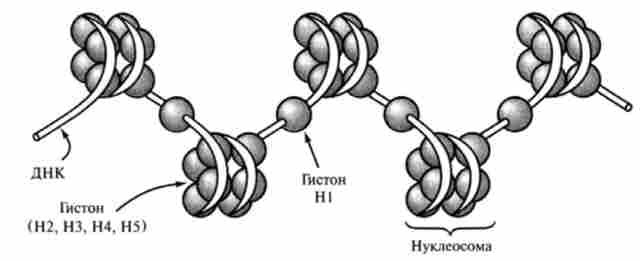
Мал. 14.6. Третинна структура ДНК еукаріот
залежності від типу клітин (від 15 до 100 нм). Лінкерних ділянки ДНК або вільні, або пов'язані з гістонів Н1, який сприяє компактизации нуклеосомної нитки і може перешкоджати транскрипції ряду генів. Гістони в клітинах піддаються ковалентного модифікації шляхомфосфорилювання, ацетилювання, метилювання і ін. Це призводить до зміни їх здатності взаємодіяти з ДНК, що є одним з механізмів регуляції транскрипції генів.
В результаті нуклеосомної організації хроматину подвійна спіраль ДНК діаметром 2 нм набуває діаметр 10-11 нм і коротшає приблизно в 7 разів.
Другим рівнем просторової організації хромосом є утворення з нуклеосомної нитки хроматиновой фібрили діаметром 20- 30 нм, що забезпечує зменшення лінійних розмірів ДНК ще в 6-7 разів. Найбільш імовірною вважається соленоидная модель упаковки в хроматиновой фібрили.
Третинний рівень організації хромосом обумовлений укладанням хроматиновой фібрили в петлі. В освіті петель беруть участь не гістонові білки, які дізналися специфічні нуклеотидні послідовності в ненуклеосомной ДНК і фіксують освіту петель. Ділянка ДНК, відповідний одній петлі, містить від 20 000 до 80 000 пар нуклеотидів і, ймовірно, представляє домен ДНК, що відповідає одиниці транскрипції. В результаті такої упаковки лінійні розміри ДНК зменшуються приблизно в 200 разів. Петлеподібна доменна організація ДНК, звана інтерфазної хромонеми, може піддаватися подальшої компактизації, ступінь якої змінюється в залежності від фази клітинного циклу (рис. 14.6).
Фізико-хімічні властивості ДНК. Різні чинники, що порушують водневі зв'язку (підвищення температури вище 80 ° С, зміна pH і іонної сили, дія сечовини і ін.), Викликають денатурацію ДНК, т. Е. Зміна просторового розташування ланцюгів ДНК без розриву ковалентних зв'язків. Подвійна спіраль ДНК при денатурації повністю або частково розділяється на складові її ланцюга. Денатурація ДНК супроводжується посиленням оптичного поглинання в УФ-області пуринових і піримідинових основ. Це явище називають гіперхромними ефектом. При денатурації зменшується також висока в'язкість, притаманна розчинів нативної ДНК. При відновленні початкової двухспіральной структури ДНК, в результаті ренатураніі, поглинання при 260 нм азотистими підставами внаслідок їх «екранування» зменшується. Це явище називають гіпохромний ефектом.
«Розплітання» кожної ДНК на складові її ланцюга здійснюється в межах певного інтервалу температур. Середня точка цього інтервалу називається температурою плавлення. Температура плавлення ДНК залежить в стандартних умовах (певна pH і іонна сила) від співвідношення азотистих основ. Г-Ц-пари, що містять три водневі зв'язки, більш міцні, тому чим більше в ДНК зміст Г- Ц-пар, тим вище температура плавлення.
Функції ДНК. У послідовності нуклеотидів в молекулах ДНК закодована генетична інформація. Основними функціями ДНК є, по-перше, забезпечення відтворення самої себе в ряді клітинних поколінь і поколінь організмів, по-друге, забезпечення синтезу білків. Ці функції ДНК обумовлені тим, що молекули ДНК служать матрицею в першому випадку для реплікації, т. Е. Копіювання інформації в дочірніх молекулах ДНК, у другому - для транскрипції, т. Е. Для перекодування інформації в структуру РНК.