| Головна |
| «« | ЗМІСТ | »» |
|---|
Хроматин, т. Е. Речовина, з якого побудована еукаріотична хромосома, являє собою складний нуклеопротеїдні комплекс. Крім молекули ДНК до складу цього комплексу входять гістонові і негістонові білки.
Гістонові білки (гістони) зустрічаються в хромосомах всіх видів еукаріотичних організмів, але відсутні у прокаріот. Вони є універсальними білками, що не володіють видовою специфічністю, і підрозділяються на п'ять основних класів, які прийнято позначати символами Я1, Н2А, Н2В, Яз і Я4.
Розрізняючи між собою за амінокислотним складом і ряду фізико-хімічних та імунологічних властивостей, гістони всіх зазначених класів є невеликими за розмірами білкові молекули, в структурі яких міститься значна кількість основних амінокислот: аргініну та лізину, - що забезпечує їх позитивний електричний заряд. Завдяки цій особливості вони можуть з'єднуватися з негативно зарядженої ДНК, утворюючи ДНК-гістонові комплекс. Таким чином, гістони виконують важливу функцію, пов'язану зі специфічною упаковкою величезною за розмірами молекули ДНК в компактну структуру хроматиди.
Негістонові білки, присутні в складі хроматину, різноманітні за структурою і виконують різні функції, що забезпечують процеси реплікації ДНК, транскрипції, відновлення пошкоджень генетичного матеріалу і ін. Ці білки можуть мати відмінності у організмів різних видів, т. Е. Характеризуватися видовою специфічністю. Вважають, що деякі негістонові білки завдяки кислотному типу їх будови (електронегативного заряду) здатні взаємодіяти з гистонами, змінюючи сумарний позитивний електричний заряд і інші властивості останніх. Такі модифікації гістонів, в свою чергу, призводять до зміни ДНК-гістонових взаємин в тих чи інших ділянках хромосоми.
ДНК-гістонові комплекси еукаріотичної хромосоми формують хроматіновие волокна, що зовні нагадують нитки бус, елементарні структурні одиниці яких отримали назву нуклеосом. Нукпеосома являє собою утворення, що складається з фрагмента ДНК величиною 140-160 пар нуклеотидів, який обвивається навколо білкового октамера (нуклеосомної кора), що має діаметр близько 10 нм і містить 8 гістонових молекул (по дві молекули кожного з гістонів: Н2А І2В, Язи // 4) (рис. 5.41).
У проміжках між окремими нуклеосомами знаходяться з'єднувальні ділянки ДНК, звані лінкерами (спейсерами), Довжина яких становить в середньому 50-60 пар нуклеотидів. З лінкерних ділянкою зв'язується молекула гистона # 1, забезпечуючи з'єднання двох сусідніх нуклеосом. Ланцюжок з'єднаних таким чином нуклеосом (полінуклео- сома) формує гвинтоподібну первинну структуру (рис. 5.41). Подальша багатоступенева укладання таких структур призводить до складної просторової їх впорядкованості, яка визначає компактну упаковку генетичного матеріалу в складі хроматину (рис. 5.42). Так, наприклад, в середню за розмірами метафазну хромосому людини, що складається з двох ідентичних хроматид довжиною близько 4 мкм, укладаються дві однакові молекули ДНК (по одній на хроматиду), кожна з яких має довжину близько 40 000 мкм (4 см).
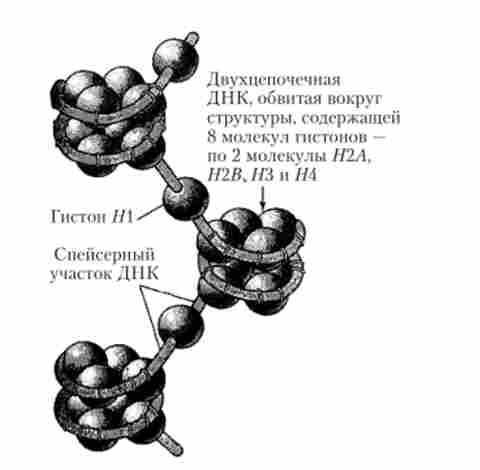
Мал. 5.41. Схематичне зображення витягнутого ділянки первинної спіралі хроматинового волокна, що складається з трьох нуклеосом
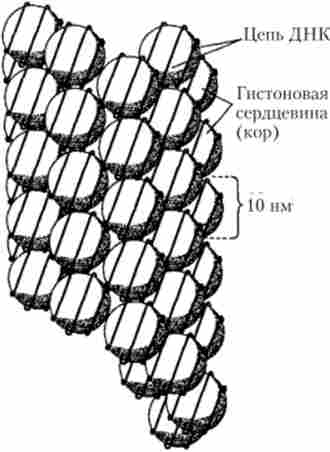
Мал. 5.42. Модель компактної упаковки нуклеосом в гвинтоподібну структуру (соленоїд), що входить до складу хроматида
Необхідно відзначити, що щільність упаковки (ступінь конденсації) хроматину неоднакова в різних ділянках однієї й тієї ж хромосоми. Щільно упакований (конденсований) хроматин отримав назву гетерахроматіна, а слабо упакований (деконденсірованний) - еухроматіна. За допомогою спеціальних методів фарбування можна диференціювати гетерохроматіновие і еухроматіновие ділянки тієї чи іншої хромосоми.
У околоцентромерних районах різних хромосом і в коротких плечах акроцентрічних хромосом присутній гетерохроматин, званий структурним (конститутивним), Який виявляється як під час мітотичного поділу клітини, так і в інтерфазних ядрі. Існує гіпотеза, що в ДНК структурного гетерохроматину або відсутні гени, які контролюють синтез метаболічно значущих білків, або в ній міститься дуже мало таких генів.
У різних хромосомах є також факультативний гетерохроматин, який виникає при деяких умовах шляхом компактизации еухроматінових районів і містить значну кількість функціонально значущих структурних генів. Однак така гетерохроматінізація того чи іншого району має оборотний характер, т. Е. Зберігається можливість повторної деконденсаціі його хроматину.
Основні дані про факультативному гетерохроматин були отримані при вивченні гігантських (політенних) інтерфазних хромосом клітин слинних залоз личинок дрозофіли, кожна з яких містить кілька тисяч копій нероздільний хроматидного структур. Є підстави вважати, що гетерохроматінізація може вибірково захоплювати різні райони хромосом всіх видів еукаріотичних організмів. Прикладом факультативного гетерохроматину у ссавців і людини є компактізацня однієї з двох Л'-хромосом в соматичних клітинах організмів жіночої статі, яка веде до формування тільця статевого хроматину інтерфазних ядер.
На відміну від еухроматінових районів гетерохроматіновие ділянки хромосом є генетично неактивними (або малоактивними), що пов'язано зі здатністю гістонів, які формують нуклеосомної кор (Н2Л, І2В, НЗ, НА), придушувати (пригнічувати) транскрипцію тісно пов'язаних з ними ділянок молекули ДНК. Отже, в гетерохроматин може інгібувати транскрипція цілої групи генів одночасно, т. Е. Виявляється принцип групової регуляції активності генів.
Очевидно, для відновлення матричної активності щільно упакованих фрагментів ДНК не потрібно повного видалення гістонові кора, але досить значного ослаблення просторової зв'язку ДНК з зазначеними гистонами (ослаблення ДНК-гістонових взаємин). Як уже зазначалося, функцію, пов'язану з деконденсація щільно упакованих районів хромосом, можуть виконувати негістонові білки регулятори, здатні змінювати електричний заряд і інші властивості гістонів. Такі білки здатні активувати роботу певних генів, дізнаючись відповідні ділянки ДНК в полінуклеосомной структурі, до яких вони специфічно прикріплюються. Передбачається, що зв'язуються з ДНК регуляторні білки дізнаються не тільки її специфічні нуклеотидні послідовності, але і ті або інші геометричні варіанти окремих ділянок цієї молекули, наприклад фрагменти молекули, що формують левозакрученной спіраль (Z-форму) ДНК.
Таким чином, уявлення про диференціальної гетерохроматі- нізації окремих районів еукаріотичної хромосоми дозволяють пояснити можливу причину розходжень в функціональної активності тих чи інших груп генів цієї хромосоми. При цьому інактивація або активація таких генів на різних етапах індивідуального розвитку організму (онтогенезу) може бути пов'язана зі змінами в структурній організації (ступеня компактизації) хроматину відповідного району хромосоми.