| Головна |
| «« | ЗМІСТ | »» |
|---|
Негативні наслідки порушення дозового балансу пов'язані з тим, що генотип є не просту суму окремих генів. Гени в генотипі об'єднані в систему завдяки складним і різноманітним взаємодій між ними, які грають важливу роль в реалізації інформації, що містяться в кожному окремому гені.
Взаємодія алельних генів. Постійно виникають різноманітні зміни структури генів обумовлюють явище множинного алелізм (див. Розд. 3.4.2.4), тому що взаємодіють при заплідненні гамети часто несуть в своїх геномах різні аллели одного і того ж гена. У зв'язку з цим генотип нового організму є гетерозиготних за багатьма локусами, т. Е. Його алельних гени, розташовані у відповідних ділянках гомологічних хромосом і прийшли від обох батьків, представлені різними алелями - А і А ', В і В', С і С 'і т. д. Якщо алельних гени представлені однаковими алелями, т. е. перебувають в гомозиготном стані (АА або А'А ', ВВ або В'В', СС або С'С '), то розвивається відповідний даному аллелю варіант ознаки. В разі Гете-розіготності (АА ', ВВ', СС ') розвиток даної ознаки (А, В або С) буде залежати від взаємодії алельних генів.
домінування - це така взаємодія алельних генів, при якому прояв одного з алелів (А) не залежить від присутності в генотипі іншого аллеля (А ') і гетерозиготи АА' фенотипично не відрізняються від гомозигот за цим алелем (АА). Така ситуація спостерігається, наприклад, коли один з алелів гена А ( «дикий») здатний забезпечити формування певного варіанту ознаки (синтез пептиду з певними властивостями), а інший А '- не володіє такою здатністю. Наявність в генотипі АА 'єдиного нормального алеля А призводить до формування нормального ознаки. Цей аллель виступає як домінантний в даному гетерозиготному генотипі. Присутність іншого аллеля (А ') фенотипично не проявляється, тому його називають рецесивним.
Прикладом домінування одного з алелей в гетерозиготному генотипі може служити визначення групової приналежності крові у людини за системою АВО. Генотипи, що містять аллель 1А або в гомозиготному стані, або в поєднанні з аллелем 1 ° (1А1А або 1А1 °), визначають розвиток у людини другої групи крові (група крові А). Така ж ситуація спостерігається і щодо алелі 1в, обумовлює формування третьої, або В-групи крові. Отже, аллели 1А і 1в виступають як домінантні по відношенню до аллелю 1 °, формує в гомозиготному стані 1 ° 1 ° першу, або 0-групу крові.
неповне домінування спостерігається, коли фенотип гетерозигот ВВ 'відрізняється від фенотипу гомозигот по обом аллелям (ВВ або В'В') проміжним проявом ознаки. Це пояснюється тим, що аллель, здатний сформувати нормальний ознака, перебуваючи в подвійній дозі у гомозиготи ВВ, проявляється сильніше, ніж в єдиній дозі у гетерозиготи ВВ '. Зазначені генотипи відрізняються експресивністю, т. е. ступенем вираженості ознаки. Демонстрацією такого типу взаємодії генів можуть бути численні спадкові захворювання у людини, які проявляються клінічно у гетерозигот по мутантом аллелям, а у гомозигот закінчуються смертю. Іноді гетерозиготи мають майже нормальний фенотип, а гомозиготи характеризуються зниженою життєздатністю.
Так, гомозиготи по аллелю серповидноклеточности еритроцитів в зв'язку з розвитком у них важкої форми анемії і інших фенотипічних проявів (див. Рис. 3.21) зазвичай не переживають дитячий вік. Навпаки, гетерозиготи - це як правило нормальні люди. Разом з тим вони все-таки відчувають кисневу недостатність в більшій мірі в порівнянні з гомозиготами по аллелю дикого типу, зокрема при підйомі на висоту (см. Розд. 4.1).
кодомінування являє собою такий тип взаємодії алельних генів, при якому кожен з алелей проявляє свою дію. В результаті цього формується якийсь проміжний варіант ознаки, новий в порівнянні з варіантами, обумовленими кожним аллелем самостійно. Прикладом може служити формування IV, або АВ-групи, крові у людини, гетерозиготного по аллелям 1А і 1в, які окремо детермінують утворення II і III груп крові.
Межаллельная комплементація відноситься до досить рідко що зустрічаються способам взаємодії алельних генів. У цьому випадку можливе формування нормального ознаки D у організму, гетерозиготного по двом мутантом аллелям гена D (D'D "). Припустимо, що ген D відповідає за синтез якогось білка, який має четвертинних структуру, що складається з декількох однакових пептидних ланцюгів. мутантний аллель D 'визначає синтез зміненого пептиду D', а мутантний аллель D "призводить до синтезу інший, але теж зміненої структури пептиду D". Можна уявити ситуацію, коли взаємодія таких змінених пептидів (D 'і D ") при формуванні четвертинної структури, ка до б взаємно компенсуючи ці зміни, забезпечує утворення білка з нормальними властивостями. У той же час окремо взаємодіючі пептиди D 'або D "формують аномальні білки. Таким чином, з певною ймовірністю у гетерозигот D'D" в результаті межаллельной комплементации може утворюватися нормальна ознака у вигляді білка з нормальними властивостями.
аллельному виняток - такий вид взаємодії алельних генів в генотипі організму, який можна зрозуміти на прикладі розглянутого вище механізму інактивації однієї з Х-хромосом у особин гомогаметний статі, що приводить у відповідність дози Х-генів у всіх представників виду. Інактивація одного з алелей в складі Х-хромосоми сприяє тому, що в різних клітинах організму, музичних по функціонуючої хромосомі, фенотипично проявляються різні аллели (рис. 3.79). Аллельному виняток спостерігається також в В-лімфоцитах, що синтезують специфічні антитіла до певних антигенів. Моноспеціфічность
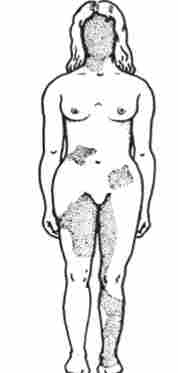
Мал. 3.79. Мозаїцизм жіночого організму за наявністю або відсутністю нормальних потових залоз в шкірі, обумовлений експресією нормального або мутантного алелей гена Х-хромосоми.
Затемнені ділянки шкіри, позбавлені потових залоз, в клітинах яких експресується мутантний аллель
таких імуноглобулінів вимагає вибору, який повинна здійснити кожна клітина між експресією батьківського або материнського алеля.
Таким чином, навіть процес формування елементарного ознаки - синтез поліпептиду з певною послідовністю амінокислот - залежить, як правило, від взаємодії щонайменше двох алельних генів, і кінцевий результат визначається конкретним поєднанням їх в генотипі.
Формування складних ознак передбачає необхідність взаємодії неалельних генів, що займають різне положення в геномі даного виду.
Взаємодія не алельних генів. Більшість ознак і властивостей організму, за якими він відрізняється від інших представників виду, є результатом дії не однієї пари алельних генів, а декількох неалельних генів або їх продуктів. Тому ці ознаки називають складними. Складним ознакою може бути четвертичная структура білка, що утворюється різними пептидними ланцюгами, які детермінуються різними (неалельних) генами. Наприклад, молекула гемоглобіну містить дві а- і дві (3-ланцюга, гени яких розташовані у людини в 16-й і 11-й хромосомах (див. Розд. 3.6.4.3). Складний ознака може бути обумовлений спільним однозначним дією декількох генів або бути кінцевим результатом ланцюга біохімічних перетворень, в яких беруть участь продукти багатьох генів. Нарешті, на формування як простих, так і складних ознак певні регуляторні впливу надають інші локуси.
Залежно від характеру участі неалельних генів у формуванні тієї чи іншої ознаки розрізняють кілька видів їх взаємодії.
Більшість кількісних ознак організмів визначається полігенами, т. е. системою неалельних генів, однаково впливають на формування даної ознаки. Взаємодія таких генів в процесі формування ознаки називають полімерним. Воно зводиться найчастіше до підсумовування дії подібних алелей цих генів, що визначають формування однакового варіанти ознаки. Спільна дія полигенов обумовлює різну експресивність - ступінь вираженості ознаки, що залежить від дози відповідних алелей. В основі появи в геномі таких генів очевидно лежить їх Дуплікація або ампліфікація (див. Розд. 3.6.4.3). Це дозволяє збільшити синтез відповідного продукту в клітинах організму.
За полімерному типу взаємодії у людини визначається інтенсивність забарвлення шкірних покривів, що залежить від рівня відкладення в клітинах пігменту меланіну. У геномі людини є чотири гени, що відповідають за цю ознаку. У генотипі всі вони представлені в подвійній дозі. У зв'язку з тим що ці гени відповідають за розвиток одного і того ж ознаки, їх позначають однією і тією ж буквою, але з різними символами, щоб підкреслити їх неал- лельность - PjP2P3P4.
Наявність в генотипі восьми домінантних алелів в системі полигенов, що визначають колір шкіри, обумовлює максимальну її пігментацію, що спостерігається в африканських негрів (Р ^ Р ^ РдРзР ^). Повна відсутність домінантних алелів у рецесивних гомозигот (р1р1р2Р2РзРзР4Р4 ^ проявляється у вигляді мінімальної пігментації у європеоїдів. Більша або менша кількість домінантних алелів, що коливається від 8 до 0, забезпечує різну інтенсивність забарвлення шкіри (рис. 3.80). Полімерне взаємодія генів лежить в основі визначення головним чином кількісних ознак (ріст, маса організму, можливо, інтелект).
Більшість складних ознак, будучи результатом цілого ланцюга біохімічних і структурних перетворень, вимагає участі багатьох генів, що впливають на різні ланки цього процесу.
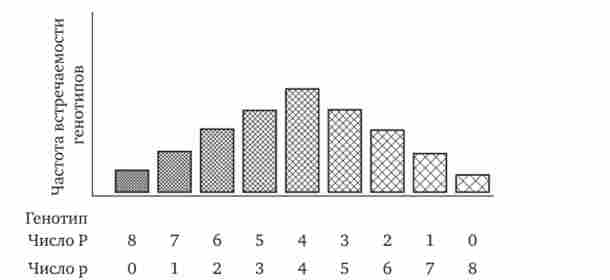
Мал. 3.80. Залежність інтенсивності пігментації шкіри у людини від кількості домінантних алелів в системі полигенов (Р) в генотипі.
Решта пояснення см. В тексті
Відсутність нормального первинного продукту, хоча б одного з них, не дозволяє сформувати нормальний складний ознака. Так як часто нормальний первинний продукт гена є результатом експресії його домінантного алеля, складний ознака формується при наявності в генотипі домінантних алелів всіх взаємодіючих генів. Така взаємодія неалельних генів, при якому вони взаємно доповнюють один одного, називають комплементарною взаємодією.
Як приклад комплементарного взаємодії неалельних генів можна розглянути процес формування статевої приналежності організму у людини. Розвиток ознак статі у людини, так само як у більшості тварин, визначається в першу чергу поєднанням гетерохромосом в його каріотипі. Наявність X- і Y-хромосом обумовлює можливість формування чоловічої статі, а двох Х-хромосом - жіночого. Однак встановлено, що для розвитку організму чоловічої статі недостатньо присутності одного лише Y-зчепленого гена, який визначає диференціювання статевих залоз за чоловічим типом і синтез ними гормону тестостерону. Для цього необхідний також продукт іншого гена - білок-рецептор, що забезпечує проникнення гормону в клітини тканин-мішеней. За синтез такого білка відповідає особливий ген, розташований в Х-хромосомі. Його мутація, що порушує утворення нормального білка-рецептора, робить тканини-мішені несприйнятливими до гормону, направляючої їх розвиток за чоловічим типом. Не використавши таку можливість на певному етапі онтогенезу, організм здійснює розвиток за жіночим типом. В результаті з'являється особина з каріотипом XY, але зовні більш схожа з жінкою. Такі суб'єкти не здатні мати потомство, так як їх статеві залози (насінники) недорозвинені, а їх вивідні протоки часто формуються за жіночим типом (недорозвинена матка, піхва). Вторинні статеві ознаки також характерні для жіночої статі. описана
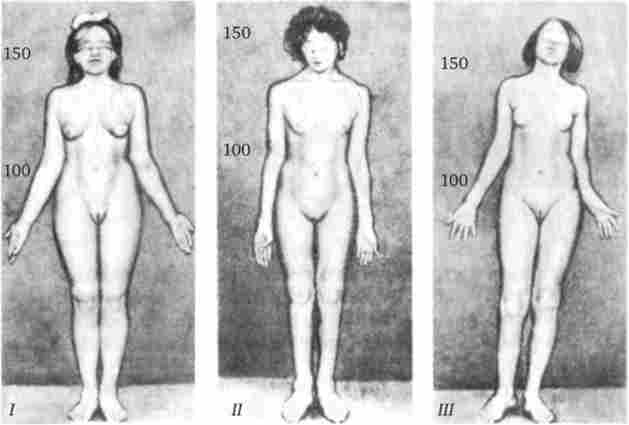
Мал. 3.81. Тестикулярная фемінізація - синдром Морріса.
I - каріотип XY (видалення сім'яників в дитячому віці); II - евнухоідний форма, каріотип XY (відсутність молочних залоз, вторинного оволосіння, природного піхви); III - каріотип XY
картина відома у людини як тестикулярная фемінізація, або синдром Морріса (Рис. 3.81).
Таким чином, досягнення кінцевого результату при формуванні у людини ознак чоловічої статі є наслідком взаємодоповнююче дії кількох (щонайменше двох) генів, що визначають можливість синтезу чоловічого статевого гормону і білка-рецептора.
Іншим прикладом взаємодії неалельних генів служить формування у людини груповий приналежності крові за системою АВО. Вище ця ознака розглядався з точки зору взаємодії алельних генів. Відповідальний за формування даної ознаки ген I забезпечує синтез антигенів А і В, що фіксуються на поверхні еритроцитів. Однак для синтезу антигенів А і В необхідно освіту неспецифічного речовини-попередника, яке детермінується геном Н, що знаходяться в домінантному стані. У гомозигот за рецесивним алелем hh навіть при наявності в генотипі домінантних алелей гена 1А або 1в антигени А і В не синтезуються і фенотипично ці люди мають I групу крові.
Така ситуація характерна для бомбейського феномена, описаного вперше в родині, де у жінки I групи крові народилася дитина з IV групою крові. Очевидно, її фенотип був обумовлений непрояв- ленням наявного в генотипі алелі 1в, що пов'язано з гомозігот- ністю матері по рецесивним аллели hh (рис. 3.82).
Ймовірно, з цих же позиції можна розглядати і випадки аль бінізма у людини, коли навіть у африканських негроїдів, кото риє володіють максимумом домінантних алелів в системі поли генів, що визначають пігментацію шкіри, з'являються альбіноси з повною відсутністю пігменту в клітинах шкіри, райдужці очей, воло сах (рис. 3.83). Альбінізм пов'язують з гомозиготностью генотипу по рецесивним аллели гена, який не належить до полігенною пігмеї тації. Можливо, його домінантний аллель визначає синтез про продукті, який бере участь в процесах, що передують синтезу

Мал. 3.83. Приклад народження альбіноса у негритянки (відсутність пігментації шкіри у дитини може бути результатом рецесивного епістазу)
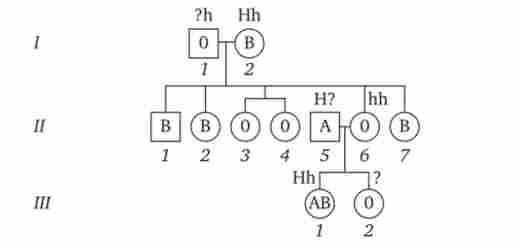
Мал. 3.82. Родовід, що ілюструє успадкування групи крові за системою АВО при бомбейському феномен.
жінка II.6 з групою крові 0 (1) має дитину III.1 з групою крові AB (IV); ймовірно, в її геномі є аллель 1в, отриманий нею від матері (1.2), але не проявився в зв'язку з її гомозиготностью по рецесивним аллели hh. Бомбейский фенотип зустрічається з частотою 1: 1 3000 серед індусів, що живуть в околицях Бомбея
пігменту. В такому випадку тут також має місце взаємодоповнююче дію відповідних алелей різних генів.
У деяких випадках при взаємодії неалельних генів для розвитку складного ознаки необхідно обов'язкову присутність одного з генів (А) в гомозиготному рецесивним стані (аа), тоді інший ген (В) забезпечує формування ознаки.
Наявність в генотипі домінантного алеля гена А якимось чином перешкоджає прояву гена В (Ь), і ознака не формується. Така взаємодія неалельних генів прийнято називати Епістатичний.
Прикладом Епістатичний взаємодії генів може бути придушення у багатьох видів розвитку забарвлення покривів, яке визначається одним з генів, при наявності в генотипі іншого гена в домінантному стані. Так, у гарбуза розвиток забарвлення плодів визначається геном В. Домінантний його аллель детермінує жовту, а рецесивний - зелене забарвлення. Однак забарвлення не розвивається взагалі, якщо в генотипі є неалельні ген А в домінантному стані. У курей породи леггорн біле оперення є результатом Епістатичний впливу домінантного алеля одного гена на прояв неалельні йому гена, відповідального за пігментацію оперення.
Відсутність необхідних даних про роль первинних продуктів багатьох генів у формуванні складних ознак часто не дозволяє точно встановити характер взаємодії неалельних локусів, що беруть участь в біохімічних процесах і складають основу утворення цих ознак. В одних випадках розвиток ознаки при наявності двох неалельних генів в домінантному стані розглядають як комплементарна взаємодія, в інших - як нерозвиненість ознаки, що визначається одним з генів при відсутності іншого гена в домінантному стані, і розцінюють як рецесивний епістаз; якщо ж ознака розвивається при відсутності домінантного алеля неалельні гена, а в його присутності не розвивається, кажуть про домінантному епістаз.
Ймовірно, саме поділ взаємодії генів на комплементарное і Епістатичний дещо штучним, бо у всіх цих випадках складний ознака є результатом поєднання в генотипі певних алелей відповідних генів, які забезпечують синтез продуктів, що беруть участь в ланцюзі біохімічних перетворень на різних рівнях формування складного ознаки.
Так як при певних поєднаннях алелей неалельних генів стає неможливим фенотипічніпрояв домінантних алелів деяких з них, нерідко спостерігається неповна пенетран- тність домінантних алелів - вони проявляються не у всіх носіїв. Так, аллели 1А і Р, що визначають групову приналежність крові за системою АВО, не проявляються фенотипно за відсутності домінантного алеля гена Н (бомбейський феномен). У альбіносів (аа) не виявляються домінантні аллели полигенов пігментації шкірних покривів (PjP2P3P4). У гарбуза не розвивається забарвлення плодів, а у курей породи леггорн - строката забарвлення оперення, контрольовані домінантними алелями відповідних генів, при наявності в їх генотипі певних неалельних генів в домінантному стані.
Особливий вид представляє взаємодія, обумовлене місцем положення гена в системі генотипу, - ефект положення. Безпосереднє оточення, в якому знаходиться ген, може позначатися на характері його експресії. Зміна активності гена, що спостерігається при хромосомних перебудовах, нерідко пов'язане з переміщенням його в іншу групу зчеплення при транслокаціях або зміною його положення в своїй хромосомі при інверсіях.
Особливий випадок, очевидно, представляє зміна експресії генів в результаті діяльності рухомих генетичних елементів, що активують або пригнічують прояв генів, поблизу яких вони вбудовуються.
Нарешті, велике значення в об'єднанні генів в єдину систему генотипу мають регуляторні взаємодії, забезпечують регуляцію генної активності. Продукти генів-регуляторів - білки-регулятори - мають здатність впізнавати певні послідовності ДНК, з'єднуватися з ними, забезпечуючи, таким чином, транскрибування інформації з структурних генів або перешкоджаючи транскрипції (див. Розд. 3.6.6).