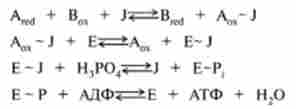
| Головна |
| «« | ЗМІСТ | »» |
|---|
При відповіді на це питання слід пояснити, по-перше, яким чином перенесення електронів служить джерелом енергії; і, по-друге, як енергія передається реакції АДФ + Р;.-? АТФ. Існує три основні гіпотези.
пояснюють механізм сполучення окислення і фосфорилювання: хімічна, хеміосмотична, конформационная.
Хімічна гіпотеза сполучення. Ця гіпотеза була запропонована більш 50 років тому. Вона постулює пряме хімічне сполучення за аналогією з субстратним фосфорилюванням. Передбачається, що існують гіпотетичні фактори сполучення (інтермедіати J), які здатні при окисленні утворювати макроергічні зв'язок (~) і потім переносити її на синтез АТФ за наведеною нижче схемою:
де А і В - переносники електронів в дихальної ланцюга; Е - фермент фосфорилювання.
В даний час ця гіпотеза досить дискредитована, оскільки багатих енергією проміжних з'єднань типу А - J не вдалося виявити.
Хеміосмотична теорія сполучення окислення і фосфорилювання. Ця гіпотеза запропонована в 1961 р П. Мітчеллом; причому значний внесок в се доказ був зроблений В. П. Скулачсвим з співавторами. Відповідно до цієї теорії, фактором, сполучаються окислення з фосфорилюванням, є електрохімічний, протонний потенціал ДРН+, що виникає на внутрішній мембрані мітохондрій в процесі транспорту електронів. При цьому передбачається, що мембрана непроникна для іонів, особливо протонів, їх гранслокація з внутрішньої сторони мембрани (з матриксу) на зовнішню сторону внутрішньої мембрани мітохондрій здійснюється за рахунок процесу окислення в дихальному ланцюгу, т. Е. Транспорту високоенергетичних електронів. Виникає електрохімічний потенціал ДРН + є адитивним; він складається з хімічного потенціалу ДРН і електричного зі знаком (+) на зовнішній стороні мембрани (Ду):
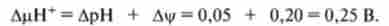
В даний час вдалося виміряти величину ДцН+, яка дорівнює 0,25 В, отже, є цілком достатньою для синтезу АТФ, при цьому більшу частину потенціалу складає Ду, т. е. електричний потенціал.
Градієнт протонів, що створює різницю хімічних і електричних потенціалів, і є джерелом енергії, необхідної для реакції
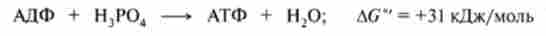
Дихальна ланцюг ферментів, утворює комплекси I, III і IV, як би тричі «перешнуровується» мембрану мітохондрій. Таким чином, кожна пара електронів, транспортують від НАДН до кисню, витягує з матриксу три пари Н+, які транслоціруется на зовнішню поверхню мембрани, в результаті чого утворюється три молекули АТФ (рис .15.7).
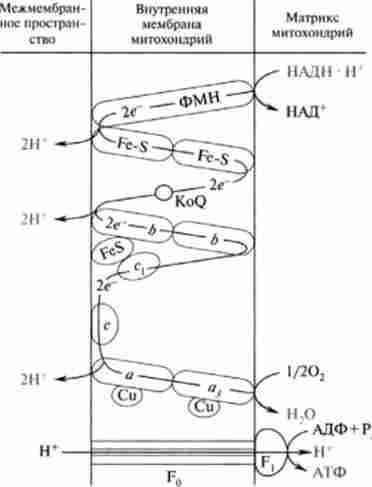
Мал. 15.7. Можлива конфігурація переносників електронів Н * вдихательная ланцюга
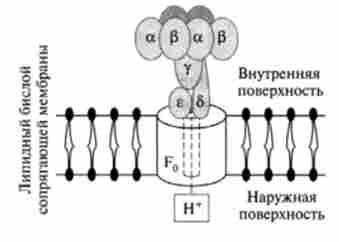
Мал. 15.8. Модель молекулярної організації Незалежної АТФ синтетазної комплексу
Яким же чином електрохімічний потенціал протонів використовується в синтезі ЛТФ? Процес фосфорилювання каталізується Незалежним АТФ-азним комплексом: Н+-АТФ-синтетаза. Цей складний комплекс складається з розчинної каталітичного компонента F, і мембранного компонента F0 (Рис. 15.8).
Компонент F, - білок з молекулярною масою 36-38 kDa - складається з п'яти типів субодиниць: а, (3, у, 6, е. Його ймовірна формула основні
каталітичні властивості компонента F, (синтез АТФ) забезпечуються а- і (3-субодиницями, у- і 8-білки здійснюють зв'язок компонента F, з іншими компонентами комплексу, а е-суб'сдініца є інгібітором АТФ-азной активності.
компонент F0 є інтегральним білком мембрани і, мабуть, наскрізь пронизує се. До складу Р0-компонента входить чотири типи субодиниць, в тому числі білок, який повідомляє даного компоненту чутливість до олігоміціну (отже, тому компонент позначається за індексом «о» - олігоміцін). компонент F0, по-перше, бере участь у зв'язуванні F, з мембраною і, по-друге, в ньому є протон-провідний канал, через який відбувається перенос Н+ із зовнішнього боку мембрани (по градієнту електрохімічного потенціалу) до компоненту F ,, який при цьому активується і стає здатним здійснити каталітичну ступінь процесу синтезу АТФ. Таким чином, так само як і комплекси дихальних ферментів, АТФ-синтетазної система фіксована в мембрані векторно, т. Е. Харастерізуется певної просторової спрямованістю, а комплекс F, виступає в матрикс і забезпечує синтез АТФ (див. Рис. 15.8).
Стадією, що лімітує синтез АТФ, є вивільнення синтезованого АТФ з активного центру ферменту в матрикс. Вважають, що енергозалежна протонирование окремих функціональних груп АТФ-азного комплексу, що відбувається за рахунок енергії ДРН+, викликає конформаційні зміни в F, -компонента, які призводять до швидкого вивільнення синтезованого АТФ з активного центру ферменту. Важливим моментом є оборотність реакції, що каталізує АТФ-азним комплексом. При відповідних умовах комплекс F0-F, може розщеплювати молекулу АТФ і використовувати отриману при цьому енергію для транспорту протонів, т. Е. Для освіти на мембрані ДРН+. Згідно з концепцією, постульованої В. П. Скулачовим, ДцН+ поряд з АТФ використовується як конвертована «валюта» для енергетичних перетворень, що протікають на мембрані. У зв'язку з цим було запропоновано всі енергетичні перетворення в клітці поділити на дві групи: що протікають в цитоплазмі (джерело енергії - АТФ, крсатінфосфат і інші макроергів) і локалізовані в мембрані, що використовують енергію ДцН+ (Рис. 15.9). Слід зазначити, що Н+ не унікальний в якості сопрягающего іона і у деяких видів організмів при певних умовах його може замінити іон натрію.
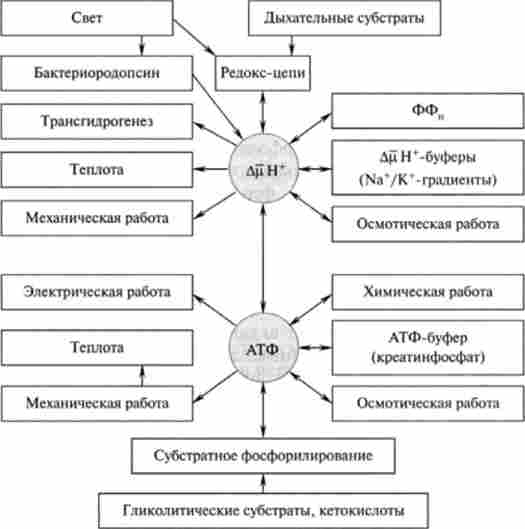
Мал. 15.9. Схема енергетики клітини, що використовує ДцН * в якості мембранної конвертованій форми енергії (по В. П. Скулачова)
Таким чином, ДцН * -залежне освіту АТФ - головний, але не єдиний процес трансформації ДцН+ в хімічну роботу. До цього ж типу енергетичних перетворень відносяться синтез неорганічного пірофосфату і перенесення відновлювальних еквівалентів в напрямку більш негативних редокс-потенціалів, наприклад зворотний перенос електронів вдихательная ланцюга і трансгідрогеназная реакція. Залежить від ДРН4 транспорт через мембрану різних речовин в бік більшої їх концентрації є трансформацію енергії за типом ДцН+ -? осмотична робота, а обертання бактеріального джгутика за рахунок енергії ДцН+ служить прикладом перетворення ДцН+ -* - механічна робота. Освіта теплоти мітохондріями тварин описується перетвореннями типу ДцН+ -> Теплопродукція.
Конфірмаціонная гіпотеза сполучення окислення і фосфорилювання. Відповідно до цієї гіпотези перенесення електронів викликає зміна конформації ліпопротеїнових ансамблів в мітохондріальної мембрани, в тому числі АТФ-синтетази, і переводить її з неактивного (релаксувати) стану в напружене, активне. Сучасні дані підтверджують різне конформационное стан крист мітохондрій при різних рівнях дихальної активності. Однак, очевидно, що пояснити активацію АТФ-синтетазної комплексу лише конформаційними переходами не представляється можливим, хоча це явище має місце.