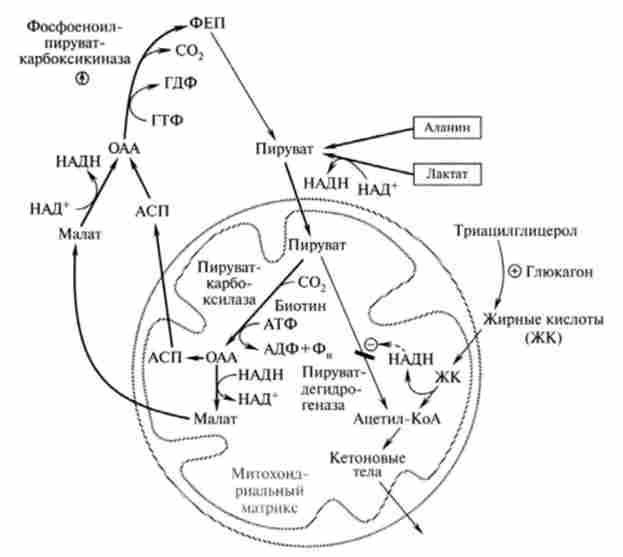
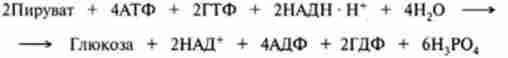Фосфорилювання пірувату. Перетворення пірувату в фосфоеноілпі- руват йде за участю двох ферментів мітохондріальної піруваткарбок- сілази (ПК) і цитозольного ферменту фосфоеноілпіруваткарбоксікінази (ФЕКК). Отже, на цій стадії процесу беруть участь два окремих субклітинних компартмента - цитозоль і мітохондрії. Схема метаболічних перетворень пірувату, в тому числі освіти фосфоеноілпі- рувата, представлена на рис. 20.3.
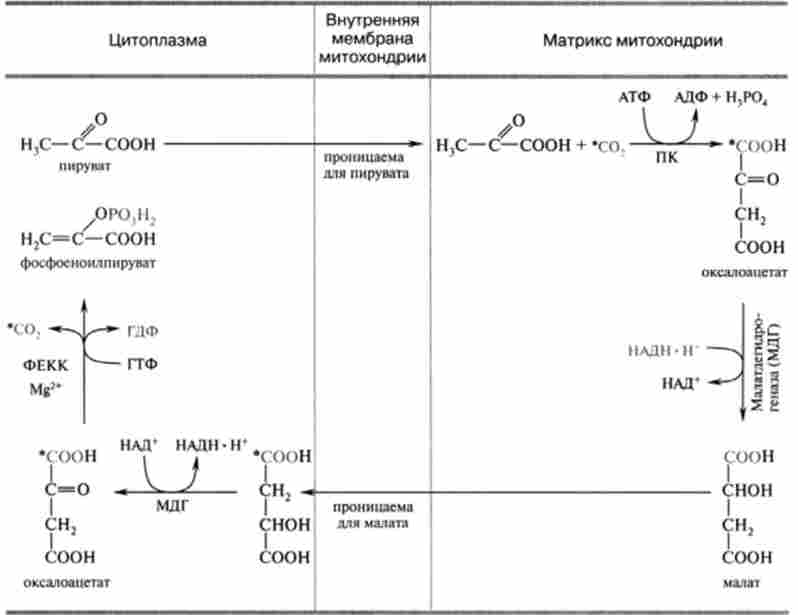
Хімізм реакції обхідного шляху фосфорилювання пірувату наведено в табл. 20.1. Перша необоротна реакція глюконеогенезу каталізується мітохондріальної піруваткарбоксілази, яка містить в якості кофермен- та вітамін Н (біотин). В мітохондріях цей фермент каталізує АТФ-зави сімую реакцію карбоксилирования пірувату, в ході якої утворюється окса- лоацетат. Для оксалоацетата внутрішня мембрана мітохондрій непроникна, і транспорт його в цитоплазму відбувається за допомогою малатного човникового механізму. Мітохондріальна малатдегідрогеназа відновлює оксало- ацетат до малата, який може виходити в цитоплазму. Потім вже цитоплазматична малатдегідрогеназа окисляє малат до оксалоацетата для подальшої участі в реакції, що каталізується фосфоеноілпіруваткарбоксікі-
назой. Продуктом цієї 1 ^2-залежною реакції, в якій донором фосфату служить ГТФ, є фосфоеноіл піруват. Зверніть увагу, що фіксований в піруваткарбоксілазной реакції С02 тепер знову відщеплюється.
Таким чином, карбоксилирование пірувату в мітохондріях мало лише енергетичне значення, і вуглець С02 (Відзначений зірочкою) в вуглецевий ланцюг продукту реакції не включається. Стехіометричне рівняння реакції:
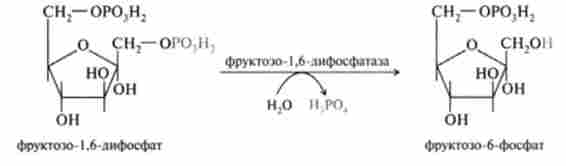
Дефосфорілірованіе фруктозо- 1,6-дифосфата і глюкозо-6-фосфату.
Ці реакції здійснюються високоспецифічними ферментами, гідроліз фосфоефірную зв'язок. Реакції є екзергонічеськие і не вимагають витрати енергії. Перетворення фруктозо-1,6-дифосфата у фруктозо-6-фосфат каталізується ферментом фруктозо-1,6-дифосфатази:
Таблиця 20.1. Механізм обхідного шляху фосфорилювання пнрувата в процесі глюконеогенезу
Цей фермент є ключовим і в кінцевому рахунку визначає здатність печінки синтезувати глюкозу з невуглеводних попередників.
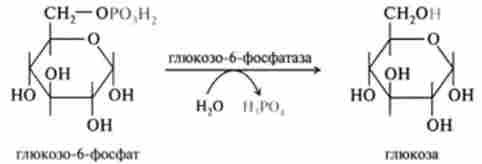
Аналогічна реакція гідролізу має місце при перетворенні глюко- зо-6-фосфату в глюкозу. Цей процес каталізується іншою специфічною фосфатазою - глюкозо-6-фосфатазою:
Глкжозо-6-фосфатаза присутній переважно в печінці і дозволяє цій тканині поставляти вільну глюкозу в кров.
Стехіометрична реакцію синтезу молекули глюкози в процесі глю- консогснеза можна записати в такий спосіб:
Таким чином, синтез глюкози з пірувату вимагає значної витрати енергії - шість високоенергетичних зв'язків (чотири від АТФ і дві від ГТФ). Крім цього, для відновних процесів потрібні ще дві молекули НАДН.
Онтогенез і патологія людини, критичні періоди в онтогенезі людини - біологія. Частина 1 З кінця XIX в. існує уявлення про наявність в онтогенетичному розвитку періодів найбільшої чутливості до шкідлива дія різноманітних факторів. Ці періоди отримали назву критичних, а фактори, що ушкоджують - тератогенних. Одностайності в оцінці різних періодів, як більш-менш стійких, не існує
Онтогенетичний рівень організації живого - біологія. Частина 1 В ієрархічній системі організації живого онтогенетичний рівень тісно пов'язаний з іншими рівнями. Елементарною одиницею життя тут є особина, в процесі її індивідуального розвитку. Реально існуючі в природі організми протягом життя безпосередньо взаємодіють з навколишнім середовищем - неживою
Оксид азоту - нервова система: анатомія, фізіологія, Нейрофармакологія Оксид азоту (NO) - маленька молекула, позбавлена заряду, добре розповсюджується дифузією, внаслідок відсутності заряду на молекулі. NO синтезується в клітинах з харчової амінокислоти аргініну під впливом ферменту NO-синтази (NOS). В даний час в організмі виявлено кілька різновидів (изоформ)
Окислення ненасичених жирних кислот - біохімія частина 2. Окислення активованих ненасичених жирних кислот (ацил-КоА) відбувається так само, як і окислення насичених кислот, т. Е. За механізмом р-окислення. Однак подвійні зв'язку природних ненасичених жирних кислот (олеїнової, лінолевої і т. Д.) Мають ^ -конфігурації, а в коа-ефірах ненасичених кислот,
Одноклітинні еукаріоти - генетика в 2 Ч. Частина 1 Світ одноклітинних еукаріот, або еукаріотичних мікроорганізмів. охоплює гриби, водорості і найпростіших. Різноманітність їх життєвих циклів і процесів, що ведуть до рекомбінації, настільки широко, а число генетично вивчених видів настільки обмежена, що великі узагальнення були б ризикованими
Обмін вуглеводів - вікова фізіологія і психофізіологія Вуглеводи складають основну частину харчового раціону і забезпечують 50-60% його енергетичної цінності. Містяться переважно в рослинних продуктах. важливо запам'ятати В організмі людини вуглеводи можуть синтезуватися з амінокислот і жирів, тому вони не відносяться до незамінних чинників харчування
Обмін речовин - анатомія центральної нервової системи У будь-якій живій клітині постійно відбуваються складні хімічні і фізичні реакції. Вони необхідні для того, щоб забезпечити постійність внутрішнього середовища як в самій клітині, так і в багатоклітинних організмі, що знаходиться під впливом мінливих зовнішніх факторів. Підтримання сталості
Обмін білків і амінокислот, загальна характеристика - біохімія частина 2. Білки є основними функціональними молекулами всіх видів живих організмів. Майже будь-яка робота в клітці - хімічна, скорочувальна, рецепторна, транспортна, імунна і багато інших виконуються білками. На відміну від вуглеводів і ліпідів білки і складові їх амінокислоти нс здатні резервуватися