| Головна |
| «« | ЗМІСТ | »» |
|---|
Зберігаючи наступність у ряді клітинних поколінь, хроматин в залежності від періоду і фази клітинного циклу змінює свою організацію. В інтерфазі при світловій мікроскопії він виявляється у вигляді грудочок, розсіяних в нуклеоплазмі ядра. При переході клітини до мітозу, особливо в метафазі, хроматин набуває вигляду добре помітних окремих інтенсивно забарвлених тілець - хромосом.
Інтерфазна і метафазну форми існування хроматину розцінюють як два полярних варіанти його структурної організації, пов'язаних в мітотичного циклу взаимопереходами. На користь такої оцінки свідчать дані електронної мікроскопії про те, що в основі як інтерфазних, так і метафазної форми лежить одна і та ж елементарна нитчатая структура. В процесі електронно-мікроскопічних і фізико-хімічних досліджень в складі интерфазного хроматину і метафазних хромосом були виявлені нитки (фібрили) діаметром 3,0-5,0,10, 20-30 нм. Корисно згадати, що діаметр подвійної спіралі ДНК становить приблизно 2 нм, діаметр нитчатой структури интерфазного хроматину дорівнює 100-200, а діаметр однієї з сестринських хроматид метафазної хромосоми - 500- 600 нм.
Найбільш поширеною є точка зору, згідно з якою хроматин (хромосома) являє собою спіралізован- ву нитку. При цьому виділяється декілька рівнів спирализации (компактизации) хроматину (табл. 3.2).
Таблиця 3.2. Послідовні рівні компактизації хроматину
|
фібрила |
ступінь укорочення |
Діаметр, нм |
|
|
в порівнянні з попередньою структурою |
в порівнянні з молекулою ДНК |
||
|
ДНК |
1 |
1 |
1-2 |
|
нуклеосомна нитка |
7 |
7 |
10 |
|
Елементарна хроматіно- вая фібрила |
6 |
42 |
20-30 |
|
інтерфазна хромонеми |
40 |
1600 |
100-200 |
|
метафазної хроматида |
5 |
8000 |
500-600 |
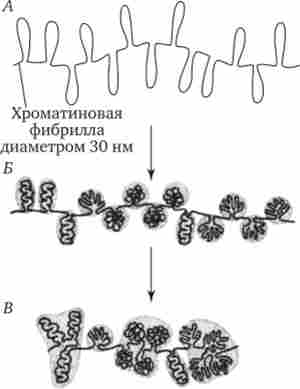
Нуклеосомна нитка - рівень організації хроматину забезпечується чотирма видами нуклеосомної гістонів: Н2А, Н2В, НЗ, Н4. Вони утворюють нагадують за формою шайбу білкові тіла - кори, що складаються з восьми молекул (по дві молекули кожного виду гістонів) (рис. 3.46).

Мал. 3.46. Нуклеосомна організація хроматину.
А - деконденсірованная форма хроматину (молекула ДНК накручена на білкові кори); Б - електронна мікрофотографія еукаріотичного хроматину (хроматин представлений нуклеосомами, з'єднаними лінкерних ДНК)
Молекула ДНК комплексируется з білковими корами, спірально накручуючи на них. При цьому в контакті з кожним кором виявляється ділянку ДНК, що складається з 146 пар нуклеотидів (п. Н.). Вільні від контакту з білковими тілами ділянки ДНК називають сполучними або лінкерних. Вони включають від 15 до 100 п. Н. (В середньому 60 п. Н.) В залежності від типу клітини.
Відрізок молекули ДНК довжиною близько 200 п. Н. разом з білковим кором становить нуклеосому. Завдяки такій організації в основі структури хроматину лежить нитка, що представляє собою ланцюжок повторюваних одиниць - нуклеосом (рис. 3.46, Б). У зв'язку з цим геном людини, що складається з 3 - 109 п. н., представлений подвійною спіраллю ДНК, упакованої в 1,5 - 107 нуклеосом.
Уздовж нуклеосомної нитки, що нагадує ланцюжок бус, є області ДНК, вільні від білкових тіл. Ці області, розташовані з інтервалами в кілька тисяч пар нуклеотидів, грають важливу роль в подальшій упаковці хроматину, так як містять нуклеотидні послідовності, специфічно впізнавані різними негістонових білками.
В результаті нуклеосомної організації хроматину подвійна спіраль ДНК діаметром 2 нм набуває діаметр 10-11 нм.
Подальша компактизація нуклеосомної нитки забезпечується гістонів HI, який, з'єднуючись з лінкерних ДНК і двома сусідніми білковими тілами, зближує їх один з одним. В результаті утворюється більш компактна структура, побудована, можливо, по типу соленоїда. Така хроматиновой фібрила, звана також елементарної, має діаметр
20-30 нм (рис. 3.47).
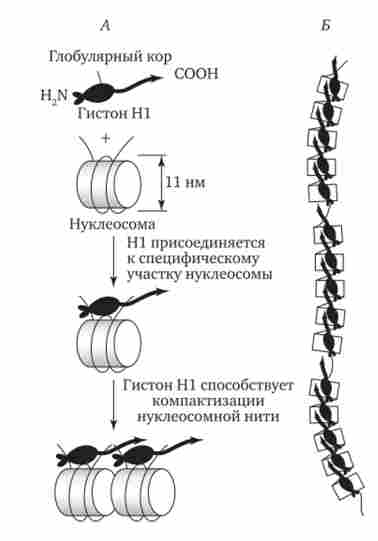

Мал. 3.47. Хроматиновой фібрила діаметром 20-30 нм.
А - з'єднання сусідніх нуклеосом за допомогою гистона HI; Б - ланцюжок, що утворюється нуклеосомами розділеними ділянками ДНК, вільними від білкових тіл; В - можлива модель упаковки ДНК в хроматіно- виття фібрили у вигляді соленоїда
Наступний рівень структурної організації генетичного матеріалу обумовлений укладанням хрома- тинів фібрили в петлі. В їх утворенні, мабуть, беруть участь негістонові білки, які здатні впізнавати специфічні нуклеотидні послідовності вненуклеосомной ДНК, віддалені один від одного на відстань в декілька тисяч пар нуклеотидів. Ці білки зближують зазначені ділянки з утворенням петель з розташованих між ними фрагментів хроматі- нової фібрили (рис. 3.48). Ділянка ДНК, відповідний одній петлі, містить від 20 000 до 80 000 п. Н. Можливо, кожна петля є функціональною одиницею генома. В результаті такої упаковки хроматиновой фібрила діаметром 20-30 нм перетворюється в структуру діаметром 100-200 нм, звану інтерфазної хромонеми.
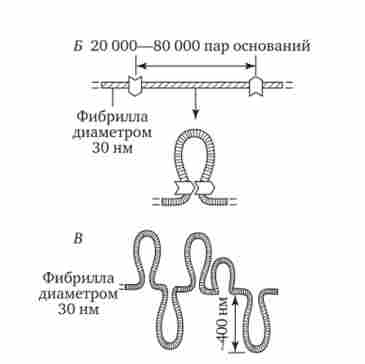
Мал. 3.48. Петельна структура хроматину - інтерфазна хромонеми.
А - хроматиновой фібрила з приєднаними до неї негістонових білками;
Б - освіту петлі на ділянці хроматіно- виття фібрили; В ?- схема петельной організації ділянки хромосоми
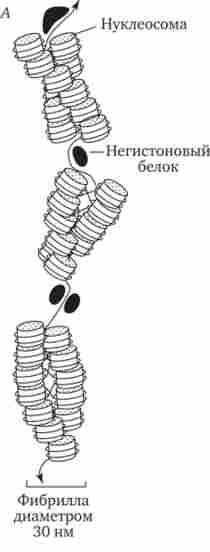
Окремі ділянки інтерфазної хромонеми піддаються подальшій компактизации, утворюючи структурні блоки, об'єднують сусідні петлі з однаковою організацією (рис. 3.49). Вони виявляються в інтерфазних ядрі у вигляді грудочок хроматину. Можливо, існування таких структурних блоків обумовлює картину нерівномірного розподілу деяких барвників в метафаз- них хромосомах, що використовують в цитогенетичних дослідженнях (див. Розд. 3.5.2.3 і 6.4.3.6).
Неоднакова ступінь компактизації різних ділянок інтерфазних хромосом має велике функціональне значення. Залежно від стану хроматину виділяють еухроматіновие ділянки хромосом, що відрізняються меншою щільністю упаковки в тиждень- трудящих клітинах і потенційно транскрібіруемих, і гетерохроматіновие ділянки, які характеризуються компактною організацією і генетичної інертністю. В їх межах транскрипції біологічної інформації не відбувається.
Розрізняють конститутивним (структурний) і факультативний гетерохроматин.
Конститутивним гетерохроматин міститься в близько-центромерних і теломерна ділянках всіх хромосом, а також протягом деяких внутрішніх фрагментів окремих хромосом (рис. 3.50). Він утворений тільки нетранскрібіруемой ДНК. Ймовірно, його роль полягає в підтримці загальної структури ядра, прикріплення хроматину до ядерної оболонці, взаємне впізнавання гомологічних хромосом в мейозі, поділі сусідніх структурних генів, участі в процесах регуляції їх активності.
Мал. 3.49. Структурні блоки в організації хроматину.
А - петельная структура хроматину;
Б - подальша конденсація хроматі- нових петель; В - об'єднання петель, що мають схожу структуру, в блоки з утворенням остаточної форми інтерфазної хромосоми
прикладом факультативного гетерохроматина служить тільце статевого хроматину, утворене в нормі в клітинах організмів гомогаметний статі (у людини гомогаметною є жіноча стать) однієї з двох Х-хромосом.
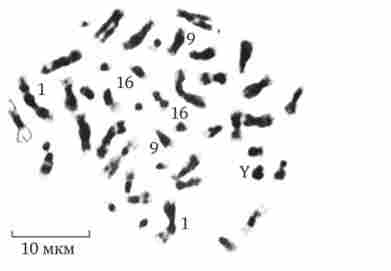
Мал. 3.50. Конститутивним гетерохроматин в метафазних хромосомах людини
Гени цієї хромосоми НЕ транскрибуються. Освіта факультативного гетерохроматину за рахунок генетичного матеріалу інших хромосом супроводжує процес клітинної диференціювання і служить механізмом виключення з активної функції груп генів, транскрипція яких не потрібно в клітинах даної спеціалізації. У зв'язку з цим малюнок хроматину ядер клітин з різних тканин і органів на гістологічних препаратах розрізняється.
Прикладом може служити Гете
рохроматізація хроматину в ядрах зрілих еритроцитів птахів.
Перераховані рівні структурної організації хроматину виявляються в клітці, що не, коли хромосоми ще недостатньо компактізовани, щоб бути видимими в світловий мікроскоп як окремі структури. Лише деякі їхні ділянки з більш високою щільністю упаковки виявляються в ядрах у вигляді хроматі- нових грудочок (рис. 3.51).

Мал. 3.51. Гетерохроматин в інтерфазних ядрі.
Компактні ділянки гетерохроматину згруповані близько ядерця і ядерної мембрани
Метафазної хромосома - вступ клітини з інтерфази в мітоз супроводжується суперкомпактізаціей хроматину. Окремі хромосоми стають добре помітні. Цей процес починається в профазі, досягаючи свого максимального вираження в метафазі мітозу і анафазе (див. Розд. 2.4.2). У телофазе мітозу відбувається декомпактізаціі речовини хромосом, яке набуває структуру интерфазного хроматину. Описана митотична супер- компактизація полегшує розподіл хромосом до полюсів митотичного веретена в анафазе мітозу.