| Головна |
| «« | ЗМІСТ |
|---|
Дія медіатора на постсинаптичну мембрану хімічного синапсу призводить до виникнення в ній постсинаптичного потенціалу. Постсинаптичні потенціали можуть бути двох типів: деполярізующіе (збуджуючі) і гіперполяризуючий (гальмівні) (рис. 5.5).
Збуджуючі постсинаптичні потенціали (ВПСП) обумовлені сумарним входять струмом позитивних зарядів всередину клітини. Такий струм може виникнути в результаті підвищення провідності мембрани для натрію, калію і, можливо, інших іонів (наприклад, кальцію).
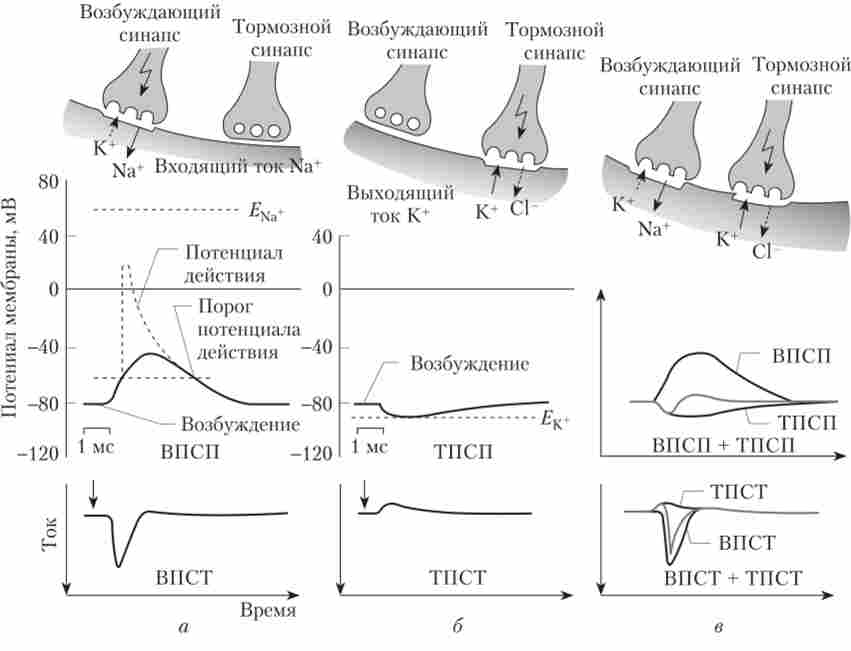
Мал. 5.5. Виникнення збуджуючих (ВПСП) і гальмівних (ТПСП) постсинаптичних потенціалів1:
а - активація тільки збуджує синапсу; б - активація тільки гальмівного синапсу; в - активація та збудливого, і гальмівного синапсів
В результаті мембранний потенціал зміщується у напрямку до нуля (стає менш негативним). Фактично величина Висі залежить від того, які іони перемістилися через мембрану і яке співвідношення проникності для цих іонів. Переміщення різних іонів відбуваються одночасно, і їх інтенсивність залежить від кількості виділився медіатора.
Таким чином, постсинаптичні потенціали є градуально реакції (їх амплітуда залежить від кількості виділився медіатора або сили стимулу). Цим вони відрізняються від потенціалу дії, який підкоряється закону «все або нічого».
Висі необхідний для генерації нервового імпульсу (ВД). Це відбувається в тому випадку, якщо Висі досягне порового значення. Після цього процеси стають незворотними, і виникає ПД. Отже, порушення в клітинах може виникати з різних причин (рис. 5.6), але в будь-якому випадку для його розвитку має відбутися зміна проникності мембрани для іонів. За аналогічним механізмам розвивається і гальмування.
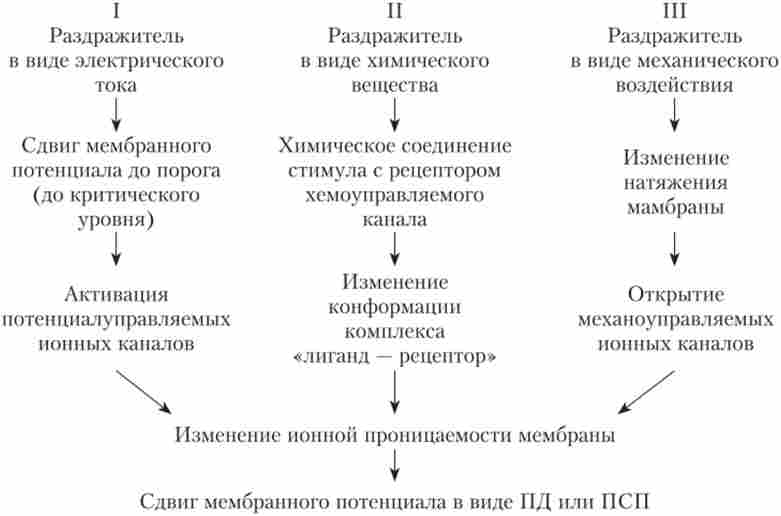
Мал. 5.6. Різні механізми розвитку збудження в клітинах
Якщо в мембрані відкриваються канали, що забезпечують сумарний виходить ток позитивних зарядів (іонів калію) або вхідний струм негативних зарядів (іонів хлору), то в клітці розвивається гальмові постсинаптичний потенціал (ТПСП). Такі струми призведуть до утримання мембранного потенціалу на рівні потенціалу спокою або до деякої гіперполяризації.
Пряме хімічне синаптичне гальмування відбувається при активації каналів для негативно заряджених іонів хлору. Стимуляція гальмівних входів викликає невелику гиперполяризацию клітини - гальмові постсинаптичний потенціал. Як медіаторів, що викликають ТГТСП, виявлені гліцин і гамма-аміномасляна кислота (ГАМК); їх рецептори пов'язані з каналами для хлору, і при взаємодії цих медіаторів зі своїми рецепторами відбувається рух іонів хлору всередину клітини і збільшення мембранного потенціалу (до -90 або -100 мВ). Цей процес називається постсинаптичне гальмування.
Однак в ряді випадків гальмування не може бути пояснено тільки в рамках постсинаптичного зміни провідності. Дж. Екклс і його співробітниками було відкрито додатковий механізм гальмування в спинному мозку ссавців: пресинаптичне гальмування. В результаті пресинаптичного гальмування відбувається зменшення вивільнення медіатора з збуджуючих закінчень. При пресінаітічному гальмуванні гальмівні аксони встановлюють синаптичний контакт з закінченнями збуджуючих аксонів. В якості медіатора пресинаптичного гальмування найчастіше зустрічається ГАМК. В результаті дії ГАМК на пресинаптичне закінчення також відбувається значне збільшення провідності для хлору і в результаті - зниження амплітуди ПД в пресинаптичному закінчення.
Функціональне значення цих двох видів гальмувань в ЦНС сильно розрізняється. Постсинаптичне гальмування зменшує збудливість всієї клітини в цілому, роблячи її менш чутливою до всіх збудливим входів. Пресинаптичне гальмування набагато більш специфічно і вибірково. Воно спрямоване на певний вхід, даючи можливість клітці інтегрувати інформацію з інших входів.