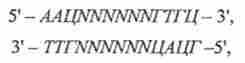
| Головна |
| «« | ЗМІСТ | »» |
|---|
Для того щоб штучним шляхом наділити будь-якої організм новими властивостями, потрібно ввести в нього новий ген або групу генів, які б там працювали - виробляли білки. Потрібний ген «в чистому вигляді» отримують декількома способами. Найчастіше його виділяють з ДНК.
Виділити ген можна, використовуючи рестрикцию ДНК, яка досягається за допомогою специфічних ферментів - рестриктаз. Рестріктази є ендонуклеази бактеріального походження, призначені для захисту клітин бактерій від чужорідної (вірусної) ДНК.
Вперше рестріктази були виявлені в клітинах кишкової палички (Е. coli), заражених бактериофагом X. При цьому виявилося, що «фагів» потомство, вирощене на двох різних штамах цієї бактерії, з різною інтенсивністю розмножується в клітинах протилежних штамів.
Так, фаг, вирощений на штамі С, погано розмножується в клітинах бактерії штаму Kl 2. Це обмеження розвитку бактеріофага пов'язано з ендонуклеазной деградацією його ДНК. Сама клітинна ДНК захищається від рестриктаз штаммоспеціфічної модифікацією - метилированием частини нуклеотидів, яке здійснюється особливим ферментом - ДНК-метилаз. Найчастіше продуктами метилування є 6-метіладенін і 5-метілцітозін.
Таким чином, присутність в клітинах бактерії двох ферментів (рестриктаз і ДНК-метилаза) забезпечує комплексний захист її ДНК. Ця система рестрикції-модифікації (A-M-система) перешкоджає схрещуванню між різними видами і штамами бактерій і тим самим забезпечує збереження їх видів в еволюції.
У генетичній інженерії рестріктази використовують для фрагментації молекул ДНК при створенні рекомбінантних геномів. Починаючи з перших робіт Д. Натанса і Г. Сміта (1962) число відкритих рестриктаз швидко росло. У 1973 р Г. Сміт і Д. Натане запропонували номенклатуру рестриктаз, яка включає наступні пункти:
При вивченні ДНК велике значення мають дві важливі особливості рестриктаз. Перша пов'язана зі здатністю ферменту дізнаватися специфічні короткі нуклеотидні послідовності в ДНК. Друга полягає в тому, що існує велика кількість різних ендонуклеаз рестрикції, кожна з яких дізнається специфічну послідовність.
Серед кількох тисяч відомих до теперішнього часу рестриктаз виділяють 3 типу.
Рестріктази типу I розривають ланцюги ДНК випадковим чином на значній відстані від ділянки впізнавання. Наприклад, сайтом впізнавання для ферменту з Е. coli К12 є
де N верхньої ланцюга - будь-яку підставу, а N нижньої ланцюга - комплементарное йому підставу.
Рестріктаза розрізає ланцюг на значній відстані від сайту впізнавання, але при цьому метилювання з утворенням 6-метіладеніна відбувається в межах цього сайту. В результаті продукти розщеплення виявляються гетерогенними, що ускладнює їх використання в генетичній інженерії.
Рестріктази типу П є основним інструментом при конструюванні рекомбінантних молекул ДНК і при аналізі структури ДНК. Ці ферменти здатні впізнавати специфічні короткі нуклеотидні послідовності, зв'язуватися з ними і робити дволанцюжкові розрізи за специфічними фосфодіестерний зв'язок або в межах самого сайту впізнавання, або на цілком певному невеликій відстані від нього. Ці розрізи можуть бути або симетричними, або несиметричними. У першому випадку утворюються так звані «тупі кінці». Так, рестриктаза нра I розрізає ДНК в тих місцях, де зустрічається послідовність 5 '... ГТТААЦ ... З'. Як і в разі інших рестриктаз типу П ця послідовність є паліндромом, т. е. в обох ланцюгах точно навпроти один одного знаходяться однакові послідовності, читаються в напрямку 5 '-> 3'.
У другому випадку дія рестриктаз призводить до утворення в ДНК так званих «липких кінців»:

З представлених схем видно, що рестриктаза нра I гидролизует фосфодіефірні зв'язку між підставами Т і А, а рестриктаза ЕсоЯ! між підставами Г і А в кожному ланцюзі. Оскільки рестриктаза розрізає обидві ланцюга в тому місці, де зустрічаються паліндромний послідовності (5 '... ГТТААЦ ... З') для нра I чи (5 '... ГААТТЦ ... З') для EcoRl, молекула ДНК розрізається на характерний для неї набір фрагментів ( «фингер- принт" - "відбитки пальців"),
рестріктаза EcoR робить ступінчасті дволанцюжкові розрізи, при цьому у утворюються фрагментів ДНК на кінцях формуються короткі комплементарні одноцепочечниє хвости з чотирьох підстав - 5'-ААТТ-3 '. При відповідних умовах комплементарні хвости возз'єднуються. Одноцепочечниє кінці, що утворюються при розщепленні ДНК реегріктазой EcoRl, отримали назву «липких», оскільки вони здатні з'єднуватися (як би злипатися) один з одним. Важливим наслідком освіти східчастих розривів є те, що фрагменти, що виходять в результаті обробки рестриктазой EcoRl двох різних ДНК (наприклад, ДНК Е. coli і дріжджів), можуть з'єднуватися за допомогою «липких кінців». При цьому відмінності, що зачіпають дволанцюжкові спіральні сегменти зазначених ДНК, на процес з'єднання не впливають.
Таке специфічне дію, що приводить до утворення певних ділянок (сайтів) рестрикції, визначило підвищений інтерес до рестріктази типу II, які стали найважливішим інструментом генетичної інженерії.
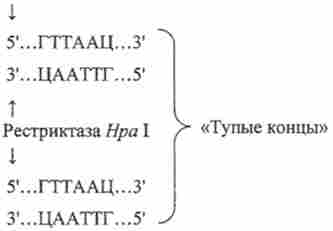
рестріктази типу II, розрізають паліндромний послідовності з утворенням «липких кінців»:

рестріктази типу II, розрізають паліндромний послідовності з утворенням «тупих кінців»:
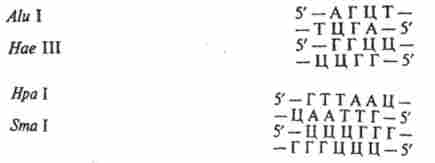
В останні роки серед рестриктаз типу II відкриті ферменти, що розпізнають непаліндромние структури і розщеплюють ДНК на строго фіксованій відстані від ділянки впізнавання з утворенням як «тупих», так і «липких кінців». Їх відносять до підкласу II S (Від англ, shift - перестановка, зрушення).
рестріктази типу II S, розрізають ланцюга ДНК на певній відстані від впізнаваною послідовності:
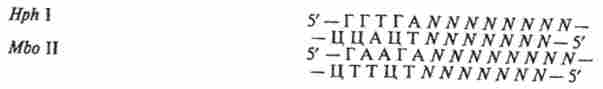
ДО 1995 року було відомо 2 400 рестриктаз типу II, що мають 188 специфічних сайтів рестрикції. Виділені з різних джерел, але мають однакову специфічність рестріктази стали називати ізоші- Мераз. Однак ізошімери не обов'язково роблять розріз в одному і тому ж місці, наприклад ХТА і Smal.
Механізм відліку такий, що місця гідролізу різних ланцюгів зміщені одна щодо іншої на один нуклеотид. В результаті утворюються фрагменти ДНК з одноланцюжковий виступами завдовжки лише в один нуклеотідний залишок.
Рестріктази типу III подібні з ферментами типу I. Вони дізнаються непаліндромние послідовності довжиною 5-6 нуклеотидних пар і розщеплюють ДНК в стороні від сайтів впізнавання на відстані 24-25 пар нуклеотидів.
У генетичній інженерії крім рестриктаз використовують і інші ферменти.
Отримання рекомбінантних молекул ДНК включає об'єднання in vitro сегментів ДНК з різних джерел. Для цього в генетичній інженерії найбільш часто використовуються ДНК-лігази, здатні зшивати фрагменти ДНК як з «липкими», так і з «тупими кінцями».
У 1958 р А. Корнбергом і його співробітниками було відкрито ДНК-полімераза Е. coli. Цей фермент, який називається нині ДНК-полімеразою I, складається з одного поліпептидного ланцюга і має трехдоменную структуру. Кожен домен має певну ферментативної активністю: // - кінцевий домен 5 '-> 3' - екзонукпеазной; С-кінцевий домен 5 '-> 3' - полімеразної (нуклеотіділтрансферазной), а середній домен 3 '-> 5' - екзонукле- азной. // - кінцевий домен може бути отщепляя з використанням протеаз (трипсину та ін.); залишається частина молекули - фрагмент Кленова - зберігає притаманну їй каталітичну активність.
Реакції, що каталізуються ДНК-полімеразою I, знайшли широке застосування. Наприклад, часто використовується здатність ДНК-полімерази I каталізувати одночасно як полімеризацію, так і 5 '- »З'-екзонуклеазное розщеплення. Фермент, виступаючи в ролі екзонуклеаза, здійснює деградацію ланцюга в 5 '- »З'-напрямку, починаючи з 5-кінця одноцепочечной проломи (ника-розриву) в двухцепочечной ДНК, а виступаючи в ролі полімерази, відновлює ланцюг шляхом послідовного приєднання мо нонуклеотідних залишків до вільної З'-гідроксильної групі на іншому кінці проломи. Власне синтезу ДНК не відбувається: пролом лише переміщається уздовж ланцюга, чим і пояснюється назва цього процесу - нік- трансляція.

Проводячи нік-трансляцію в присутності 32Р-мічених нуклеотидів, в якості субстратів отримують мічені фрагменти ДНК з високою питомою активністю. ця реакція in vivo відіграє важливу роль у репарації пошкодженої ДНК.
У 1964 р Г. Темін висунув гіпотезу про існування специфічних для РНК-містять ретровірусів ферментів, здатних синтезувати ДНК на матриці РНК. У 1970 р Г. Темін і С. Мізутані і незалежно від них Д. Балтімор відкрили цей фермент у вірусу саркоми Рауса. Ця РНК-залежна ДНК-полімераза отримала назву зворотної транскриптази, або ревертази. Найбільш вивчена ревертаза вірусів птиці, яка складається з 2 субодиниць - а й р - і володіє принаймні трьома активностями: ДНК-полімеразної (може використовувати в якості матриці як РНК, так і ДНК); активністю РНКази Н (гідролізує РНК у складі гібрида РНК: ДНК, але не атакує вільну РНК); ДНК-ендонуклеазной. Перші дві активності використовуються для синтезу ДНК, комплементарної вірусної РНК, яка потім інтегрується в геном клітини-господаря. Ендонуклеазная активність, очевидно, використовується для внесення розривів в ланцюзі
ДНК-господаря. Встановлено, що Р-субодиниці даної ревертази володіють усіма трьома активностями, а-субодиниця - тільки активностями ДНК-полімерази і РНКази Н. затравки (праймером) для полімеразної реакції можуть служити невеликі ділянки одноланцюгових молекул ДНК або РНК (а також тРНК).
У генетичній інженерії ревертаза широко використовується для цілеспрямованого синтезу на матричних РНК комплементарних молекул ДНК (кДНК). Для підвищення ефективності цієї процедури додатково застосовують і інші ферменти (рис. 15.1).
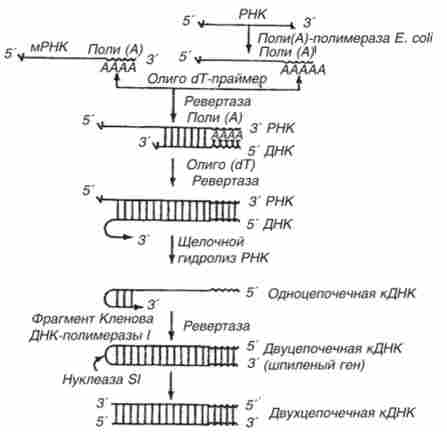
Мал. 15.1. Схема отримання кДНК з використанням ревертази вірусу пташиного міелобластоза і трьох додаткових ферментів: полі (А) -полімерази, фрагмента Кленова ДНК-полімерази I і нуклсази Я (руйнує характерну шпильку в молекулі кДНК, яку утворює ревертаза)
Важливе значення в експериментах з рекомбінантними ДНК має полі (А) -полімераза, яка приєднує нуклеотидні залишки до -кінцю ланцюга без участі матриці. Полі (А) -полімераза виконує високоспеціалізовану функцію в експериментах з рекомбінантними ДНК: її використовують для приєднання полі (А)-кінців до молекул РНК при синтезі кДНК.