| Головна |
| «« | ЗМІСТ |
|---|
Основні функції нейрона - генерація, проведення та передача потенціалу дії - стають можливі, так як між його зовнішньої і внутрішньої середовищем існує різниця концентрацій іонів калію (К +), натрію (Na +), кальцію (Са2 +) і (С1). У внутрішньому середовищі більше К + і менше Na +, Са2 + і С1 ~, а в зовнішньому середовищі навпаки. Така різниця концентрацій підтримується роботою іонних білків-насосів, вбудованих в цитоплазматичну мембрану. Наприклад, Иа + -К + -АТФаза (натрій калієвий насос) постійно переносить катіони (позитивно заряджені частинки) калію всередину клітини і виносить з клітки катіони Na + (активний транспорт). В результаті концентрація К + в клітці приблизно в 30-40 разів більше, a Na + - в 10-12 разів менше, ніж в міжклітинної рідини.
У мембрані клітини присутні також численні білки-канали, які можуть вибірково пропускати через себе різні іони. Частина цих білків має постійно відкриті канали, інша частина має канали зі стулками, що відкривають канал тільки за певних умов і таким чином регулюють ноток іонів через мембрану. Через канали іони пересуваються в результаті дифузії - пересування частинок з області з більшою концентрацією в область з меншою. Отже, через калієві канали К + виходить з клітки, а через натрієві, кальцієві і хлорні відповідні іони в клітку входять. Пересування іонів через канали не вимагає витрат енергії клітиною (пасивний транспорт).
Завдяки нерівномірного концентрації іонів мембрана нейрона поляризована, т. Е. Між внутрішньою і зовнішньою її поверхнею існує
різниця потенціалів - мембранний потенціал (МП), причому більшу частину часу внутрішня поверхня заряджена негативно. Збільшення поляризації мембрани називають гиперполяризацией (внутрішній негативний заряд збільшується), а зменшення поляризації - деполяризацией. Негативний заряд внутрішньої поверхні мембрани характерний для будь-яких клітин організму, і у більшості клітин він підтримує одну й ту величину протягом усього життя. Але в клітинах збудливих тканин (нервової, м'язової, залізистої) його величина може змінюватися, тому в таких клітинах МП в стані фізіологічного спокою клітини називається потенціалом спокою (ПП). Середня його величина для нейрона становить -70 мВ. ПП генерується завдяки перерозподілу іонів через постійно відкриті калієві і натрієві канали. При різних впливах на нейрон відбувається перерозподіл іонів, в результаті чого генеруються потенціали дії (ПД, нервові імпульси), постсинаптичні потенціали, рецепторні потенціали.
Розглянемо більш докладно, як генерується ПД. Він являє собою швидке коливання мембранного потенціалу, що супроводжується перезарядженням мембрани (рис. 2.17), т. Е. Усередині вона стає зарядженої позитивно, а зовні - негативно.
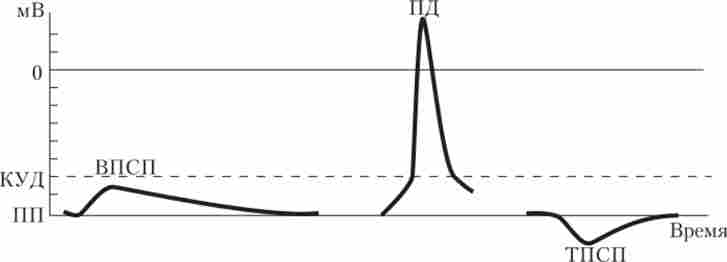
Мал. 2.17. Електричні потенціали нервової клітини:
ПП - потенціал спокою; КУД - критичний рівень деполяризації; ПД - потенціал дії; У ПСП - збудливий постсинаптичний потенціал;
Тисп - гальмівний постсинаптичний потенціал
Для генерації ПД необхідні потенціалзалежні Na+- і К+-канали. Вони відкриваються, якщо мембрана деполяризуется до порогової величини, т. Е. До такого рівня потенціалу, який призводить до генерації ПД. Таку величину називають критичним рівнем деполяризації (КУД). В середньому порогова деполяризація дорівнює -50 мВ. Першими і дуже швидко відкриваються Иа+-канали, що викликає стрімкий потік катіонів натрію в клітку і зменшення поляризації мембрани. Натрію входить так багато, що заряд в клітці стає позитивним, досягаючи значень від +20 до +50 мВ. Коли ПД досягає свого піку, канали закриваються (инактивируются). Таким чином, вхід натрію забезпечує висхідну фазу 11Д. Калієві канали відкриваються набагато повільніше натрієвих, і виходить ток калію досягає максимуму тільки після ника ПД, коли натрієві канали вже закриті. За рахунок виходу К+ позитивний заряд в клітці стає все менше (реполяризация), і поступово заряд мембрани досягає рівня IIII. Таким чином, вихід калію забезпечує спадну фазу Г1Д.
Особливістю ПД є те, що при пороговому подразненні він відразу досягає своєї максимальної амплітуди і збільшення подразника до сверхпороговое значень не змінює величину ПД. Цю особливість називають законом «все або нічого». Так як ПД є основним носієм інформації в НС, стає зрозумілим, що кодувати її величиною ПД неможливо, тому для кодування в першу чергу використовується частота генерації ПД нейроном.
Після генерації ПД починається поширення його по мембрані. Розглянемо, як відбувається цей процес в безмиелиновом волокні. Коли ПД генерується, наприклад, в одній з точок аксона, це призводить до входу в волокно великої кількості Na+, дифузії його в сусідні ділянки і деполяризації сусідніх ділянок, що досягають порогового рівня. В результаті відкриваються розташовані на цих ділянках потенціалзалежні Na+- і К+-Капали, що призводить до генерації чергового ПД і т. д. В мієлінових волокнах потенціалзалежні канали присутні тільки в перехоплення Ранвей, в результаті чого ПД генеруються тільки в перехоплення, що і збільшує швидкість поширення ПД в цих волокнах. Принципово важливо, що кожен знову генерується IIД має одну і ту ж амплітуду, т. Е. поширюється по волокну без загасання.
Коли ПД, поширюючись по волокну, доходить до пресинаптичного закінчення, це викликає виділення медіатора в синаптичну щілину. Медіатор поширюється в ній, доходить до білків-рецепторів, що сидять на постсинаптичні мембрані, і контактує з ними за принципом «ключ до замка». Контакт медіатора і рецептора призводить до відкривання хемозавісімих (ліганд-залежних) каналів, потоку іонів через ці канали і генерації постсинаптичного потенціалу (ПСП). На відміну від ПД, ПСП градуально, т. Е. Їх величина може змінюватися в залежності від кількості виділився з пресинапса медіатора. Величина ПСП значно менше, ніж ПД, всього 5-10 мВ. Крім того, ці потенціали, поширюючись по мембрані від місця свого виникнення, загасають, т. Е. Не можуть, як ПД, поширюватися на великі відстані.
Характер ПСП залежить від того, які іони будуть проходити через канали. Якщо на постсинаптичні мембрані присутні Na+- і (або) Са++-канали, то що входять через них позитивні іони зрушать ПП вгору (деполярізуют мембрану). Потенціал мембрани стане ближче до рівня порогової деполяризації, т. Е. Клітина стане щодо більш збудливою. Тому такий потенціал називають збудливим постсинаптичним потенціалом (ВПСП). Синапси, в яких генеруються ВПСП, називають збудливими. І навпаки, якщо відкриваються До+- або сь-канали, то вихід К+ або вхід С1_ призводять до гіперполяризації (зрушенню потенціалу до більш негативних значень), що робить клітину щодо загальмованою. Такий потенціал називають гальмівним ПСП (ТПСП), і синапси, в яких генеруються ТПСП, називають гальмівними.
Як вже говорилося, на мембрані одного нейрона знаходяться тисячі синапсів. Приблизно половина з них - збуджуючі. Для того щоб постсинаптичні нейрон згенерував новий ПД і таким чином передав інформацію далі по мережі нейронів, потрібно, щоб його мембрана досягла рівня порогової деполяризації. Але величина одного ВПСП цього рівня не досягає, необхідна сумація декількох ВПСП. При цьому треба враховувати, що одночасно з ВПСП в гальмівних синапсах генеруються ТПСП. У різні періоди часу більш-менш активні або збуджуючі, або гальмівні синапси. І якщо збуджуючі синапси активніші і сумарна величина всіх збудливих і гальмівних ностсінаптічних потенціалів досягне порогового рівня, то нейрон згенерує новий ПД, і поширення інформації продовжиться. Якщо ж більш активні гальмівні синапси, то нейрон буде перебувати в загальмованому стані, і генерація їм ПД буде можлива тільки при дії подразника більшої сили.
Такі основні закономірності роботи будь-якого нейрона як з мільярдів нейронів нашої НС, так і з десятків нейронів найпростіших НС безхребетних тварин.