| Головна |
| «« | ЗМІСТ | »» |
|---|
Вище було показано, що експерименти по виявленню термінів детермінації в ранньому розвитку різних видів тварин привели до відкриття явища ембріональної регуляції, т. Е. Процесу відновлення нормального розвитку цілого зародка або зачатка після його природного або штучного порушення. Досліди, численні і дуже різноманітні, дали цікаві результати.
Для вивчення регуляційних можливостей зародків використовували такі методичні прийоми: 1) видалення частини матеріалу зародка; 2) додавання надлишкового матеріалу; 3) перемішування матеріалу, а також зміна просторового взаєморозташування шляхом центрифугування або здавлення. Ці маніпуляції проводили на стадіях яйця, зиготи, дроблення, гаструляції і органогенезу.
До початку дроблення від яйцеклітин відбирали частину цитоплазми шляхом відсмоктування піпеткою або відриву в результаті сильного центрифугування. Виявлено, що за дуже рідкісними винятками вилучення навіть 1/3 цитоплазми яйця не перешкоджає подальшому нормальному розвитку зародка, хоча і меншого розміру. Такі результати були отримані на яйцеклітинах кишковопорожнинних, многоще- Тинкову черв'яків, молюсків, асцидій, голкошкірих. Злиття двох або декількох яйцеклітин теж закінчувалося нормальним розвитком.
Досліди по центрифугированию яєць теж дали цікаві результати. Показано, що сильне центрифугування яєць молюсків, черв'яків і морського їжака не змінює місця виділення полярних тілець, напрямки першої борозни дроблення і характеру розвитку. Виходить абсолютно нормальна личинка з випадковим розподілом пігменту і інших речовин. Яйце тритона після 20-хвилинного центрифугування теж дробиться, незважаючи на повне порушення розташування в ньому жовтка і інших компонентів. Існує припущення, що певне розміщення різних сортів цитоплазми всередині яйця і зиготи не їсти причина всього подальшого розвитку, а, навпаки, саме просторове розташування цитоплазми є наслідком інших, більш глибоких причин, які потрібно шукати в енергетиці та фізіології яйця.
Всі наведені досліди показують, що овоплазматічна сегрегація НЕ є абсолютною причиною детермінації і при її порушенні можлива ембріональна регуляція.
Безліч дослідів проведено в стадії дроблення. Майже у всіх видів тварин поділ бластомерів на перших етапах дроблення призводить до розвитку з них цілих, нормальних зародків, т. Е. Моно- зиготних близнюків. У морського їжака кожен з чотирьох бластомерів може утворити нормальну личинку. На стадії восьми бластомерів повноцінна регуляція здійснюється в бластомерах вегетативного полюса, а в бластомерах анімального полюса вона можлива при приміщенні їх в слабкий розчин солей літію.
Регуляція під час дроблення широко поширена практично у хребетних: міног, хрящових і кісткових риб, земноводних і вищих хребетних. Про це свідчать досліди по ізоляції бластомерів і часте утворення монозиготних близнюків і двойнікових
каліцтв. Можливий розвиток нормальних кроленят з одного віддаленого бластомера двох-, чотирьох- і восьміклеточной стадії зародка в 30,19 і 11% випадків відповідно. У четирехклеточного зародка миші при поділі бластомеров можливий розвиток трьох бластоцист і одного трофобластичного бульбашки. З цих дослідів випливає, що потенції окремого бластомера до морфогенезу ширше, ніж реалізуються з нього в ході нормального розвитку зачатки. потенції - це максимальні можливості елементів зародка, т. е. напрямки їх розвитку, які могли б здійснитися. У нормі реалізується лише одне з них, а інші можуть бути виявлені в експерименті. Широкі потенції називають ще тотипотентностью.
Широкі потенції бластомеров підтверджуються численними дослідами по їх пересадці і перемішування. Бластомери морського їжака поділяли і потім перемішували безладно. Вийшли нормальні личинки, але іншим шляхом: кишечник, наприклад, утворювався НЕ инвагинацией, а шізоцельно (шляхом розбіжності) з щільної клітинної маси; скелет виникав раніше, ніж покриви. Це приклад еквіфінал'ності, яку можна розглядати як одне з найважливіших властивостей регуляційних процесів.
У зародків ссавців бластомери при пересадці поводяться відповідно до місця пересадки. Це підтверджено перенесенням клітин з трофобласта в зародковий вузлик, і навпаки. У цих випадках виявляються залежність долі бластомера від взаємодії з іншими бластомерами і ширші його потенції. У такому ж ключі можна інтерпретувати такі досліди: якщо бластоцисту помістити в порожнину тіла, то з неї розвивається солідна пухлина (ембріокарцінома). Клітини цієї пухлини, будучи введені в бластоцисту, призводять до нормального розвитку зародка.
Великий інтерес представляють досліди по об'єднанню зародків. Штучне злиття двох зародків морського їжака на стадії бластули може завершитися формуванням абсолютно нормальної структури личинки. Така личинка перевершує за розмірами нормальних зародків і має вдвічі більше клітин. Це доводить, що при абсолютно тотожною архітектоніці тіла воно може бути побудовано з різної кількості клітин. Отже, детермінація носить не клітинний характер, а є похідною від організму в цілому.
Відносно ссавців було висловлено припущення, що в їх зародках предетермінірованние локалізовані ділянки цитоплазми не грають ніякої ролі. Залежність долі клітини від її положення була продемонстрована в такий спосіб. Об'єднували клітини двох мишачих зародків, які перебувають на стадії морули і попередньо дисоційованому. Зародки відрізнялися по генам забарвлення шерсті. Що утворилася в результаті гібридну бластоцисту імплантували в прийомну матір. Розвинувся нормальний живий мишеня-химера, в забарвленні якого проявилося дію обох генів обох батьків (рис. 8.19).
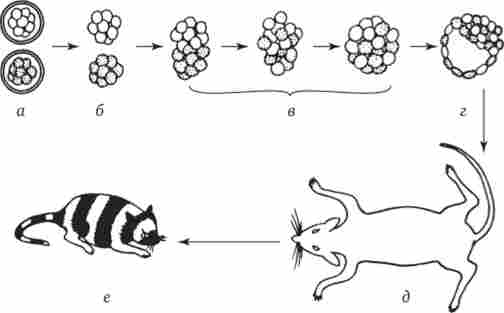
Мал. 8.19. Химерний мишеня, отриманий в результаті об'єднання на стадії морули двох зародків з різними генотипами.
а - дробящиеся яйця, 6 - морули, звільнені від яйцевих оболонок шляхом обробки протеолітичнимиферментами, в - об'єднуються морули, інкубовані in vitro при 37 ° С, г - об'єднаний зародок на стадії бластоцисти, д - мати-реципієнт з імплантованим химерним зародком, е - химерний мишеня
Наявність періоду розвитку, коли потенції елементів зародка ширше, ніж реалізуються при нормальному розвитку, т. Е. Коли детермінація не носить остаточного, необоротного характеру, лежить в основі ембріональних регуляцій. Ембріональні регуляції були відкриті німецьким ембріологів Г. Дріш (1908). Таким чином, детермінація і ембріональна регуляція є протилежними властивостями і найтіснішим чином пов'язані між собою в системі цілісного зародка.
Детермінація, як правило, йде від цілого до частин: спочатку детермінується цілий зачаток зародка, але доля окремих його елементів (клітин) ще не визначена, потім поступово або стрибкоподібно детермінуються окремі елементи. Зазвичай детермінація змінюється диференціацією і спеціалізацією частин організму, посилюється взаємна залежність їх один від одного, зростають інтегрованість, цілісність організму. Виняток становлять види, у яких дорослі форми влаштовані простіше личинок. Здатність до ембріональної регуляції в ході онтогенезу відповідно падає, але не абсолютно, так як і у дорослого організму існує, наприклад, здатність до регенерації, тобто. Е. До відновлення втрачених або пошкоджених частин.
Важливо, що не тільки в експерименті, де штучно порушується нормальний хід розвитку, але і в природних умовах хід розвитку клітин регулюється відповідно до їх положенню в цілому.
Ймовірно, гнучкий регуляторний характер розвитку має суттєві адаптивні переваги: якщо якесь одна ланка буде придушене або повністю випадає, зберігаються шанси
на досягнення нормального кінцевого результату. З іншого боку, в екстремальних умовах резерв мінливості може стати джерелом еволюційних перетворень. Ці припущення цілком узгоджуються з фактом регуляційного типу яєць у хребетних і з тим, що взагалі здатність до ембріональної регуляції у них виражена сильніше, ніж у багатьох безхребетних.