| Головна |
| «« | ЗМІСТ | »» |
|---|
морфогенез - це процес виникнення нових структур і зміни їх форми в ході індивідуального розвитку організмів. Морфогенез, як зростання і клітинне диференціювання, відноситься до ациклічним процесам, т. е. які повертаються до свого попереднього стану і здебільшого незворотнім. Головною властивістю ацикличних процесів є їх просторово-часова організація. Морфогенез на надклеточном рівні починається з гаструляції. У хордових тварин після гаструляції відбувається закладка осьових органів. У цей період, як і під час гаструляції, морфологічні перебудови охоплюють весь зародок. Наступні потім органогенезу є місцеві процеси. Усередині кожного з них відбувається розчленовування на нові дискретні (окремі) зачатки. Так послідовно в часі і в просторі протікає індивідуальний розвиток, що приводить до формування особини зі складною будовою і значно багатшою інформацією, ніж генетична інформація зиготи.
Морфогенез пов'язаний з дуже багатьма процесами, починаючи з прогенезу. Поляризація яйцеклітини, овоплазматічна сегрегація після запліднення, закономірно орієнтовані ділення дроблення, руху клітинних мас в ході гаструляції і закладок різних органів, зміни пропорцій тіла - все це процеси, що мають велике значення для морфогенезу. Крім надклеточних рівня до морфопроцессам відносяться такі процеси, які протікають на субклітинному і молекулярному рівнях. Це зміни форми і будови окремих клітин, розпад і відтворення молекул і великих молекулярних комплексів, зміна конформації молекул.
Таким чином, морфогенез являє собою багаторівневий динамічний процес. В даний час вже багато відомо про тих структурних перетвореннях, які відбуваються на внутрішньоклітинному і міжклітинному рівнях і які перетворять хімічну енергію клітин в механічну, т. Е. Про елементарні рушійні сили морфогенезу.
У розшифровці всіх цих внутріуровневие і міжрівневих процесів велику роль зіграв каузально-аналітичний (Від лат. Causa - причина) підхід. Даний відрізок розвитку вважають поясненим, якщо його вдалося представити у вигляді однозначної послідовності причин і наслідків. В цьому аспекті одним з першорядних є питання про те, чи міститься в геномі даного виду або в генотипі зиготи інформація про конкретних морфологічних процесах. Очевидно, що в геномі даного виду закладена інформація про кінцевий результат, т. Е. Розвитку особини певного виду. Очевидно також, що в генотипі зиготи містяться певні аллели батьків, що володіють можливістю реалізуватися в певні ознаки. Але з яких саме клітин, в якому місці і в який конкретно формі розвинеться той чи інший орган, в генотипі НЕ закладено.
Це твердження випливає з усіх відомостей про явища ембріональної регуляції, які показують, що конкретні шляхи морфогенезу як в експерименті, так і в нормальному розвитку можуть варіюватися. гени, позбавлені однозначного морфогенетичного сенсу, набувають його, проте, в системі цілісного що розвивається організму і в контексті певних, структурно стійких схем морфогенезу.
Клітини і клітинні комплекси роблять закономірні спонтанні, що не породжуються зовнішніми силами, макроскопічні морфогенетичні руху. При зміні положення, зменшенні або збільшенні кількості бластомерів і при пересадці ембріональних індукторів в нетипове місце нерідко досягається нормальний результат. Це дозволяє розглядати морфогенез як самоорганізується процес утворення структур з початково однорідного стану, що є невід'ємною властивістю систем, що самоорганізуються, що володіють властивістю цілісності.
Одночасно з взаємозв'язком всіх частин ембріона виникають щодо автономізованих біологічні системи, здатні продовжувати розвиток в умовах ізоляції від цілого організму. Якщо зачаток стегна курячого зародка культивувати в штучному середовищі, він продовжує розвиватися в колишньому напрямі. Око щури, ізольований на стадії 14-17 діб, продовжує автоматично розвиватися, хоча дефектно і повільніше. Через 21 добу очей в культурі тканин набуває ту ступінь складності структури, яку нормально він вже має на 8-у добу після народження щури. Для пояснення всіх цих явищ каузально-аналітичний підхід непридатний. На озброєння взята фізико-математична теорія самоорганізації нерівноважних природних систем, як біологічних, так і синтетичних.
Концепція фізіологічних градієнтів, запропонована на початку XX ст. американським вченим Ч. Чайльд, полягає в тому, що у багатьох тварин виявляються градієнти інтенсивності обміну речовин і збігаються з ними градієнти пошкоджуваності тканин. Ці градієнти зазвичай знижуються від переднього полюса тваринного до заднього. Вони визначають просторове розташування морфогенезу і цітодіфференціровкі. Виникнення самих градієнтів визначається гетерогенністю зовнішнього середовища, наприклад поживних речовин, концентрації кисню або сили тяжіння. Будь-яке з умов або їх сукупність можуть викликати первинний фізіологічний градієнт в яйцеклітині. потім можливо
виникнення вторинного градієнта під деяким кутом до першого. Система з двох градієнтів (або більше) створює певну координатну систему. Функцією координати є доля клітини.
Більш сучасною є концепція позиційної інформації, по якій клітина як би оцінює своє місце розташування в системі координат зачатка органу, а потім диференціюється відповідно до цього положення. На думку сучасного англійського біолога Л. Вольперт, положення клітини визначається концентрацією деяких речовин, розташованих уздовж осі зародка за певним градієнтом. Відповідь клітини на своє місце розташування залежить від генома і всієї попередньої історії її розвитку. На думку інших дослідників, позиційна інформація є функція полярних координат клітини. Існує також думка про те, що градієнти є стійкими сліди періодичних процесів, що розповсюджуються вздовж розвивається зачатка. Концепція позиційної інформації дозволяє формально інтерпретувати деякі закономірності онтогенетичного розвитку, але вона дуже далека від загальної теорії цілісності.
концепція морфогенетичних полів, базується на припущенні про дистантних або контактних взаємодіях між клітинами зародка, розглядає ембріональний формоутворення як самоорганізується і самоконтрольованого процес. Попередня форма зачатка визначає характерні риси його подальшої форми. Крім того, форма і структура зачатка здатні надати зворотну дію на біохімічні процеси в його клітинах. Найбільш послідовно цю концепцію розробляв в 1920-1930-х рр. вітчизняний біолог А. Г. Гурвич, який запропонував вперше в світовій літературі математичні моделі формоутворення. Він, наприклад, моделював перехід ембріонального головного мозку зі стадії одного міхура в стадію трьох бульбашок.
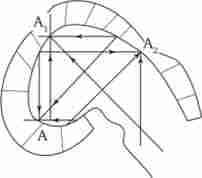
Мал. 8.20. Моделювання морфогенезу головного мозку зародка курки
Модель виходила з гіпотези про відразливих взаємодіях між протилежними стінками зачатка. На рис. 8.20 ці взаємодії відображені трьома векторами (A, Av Л2). Гурвич вперше вказав також на важливу роль нерівноважних надмолекулярних структур, характер і функціонування яких визначаються доданими до них векторами поля. В останні роки К. Уоддінгтон створив більш узагальнену концепцію морфогенетичного векторного поля, що включає не тільки формоутворення, але і будь-які зміни, що розвиваються.
Близько ідеї лежать в основі концепції дисипативних структур.
Диссипативними (від лат. Dissipatio - розсіювання) називають енергетично відкриті, термодинамічно нерівноважні біологічні і небіологічні системи, в яких частина енергії, що надходить в них ззовні, розсіюється. В даний час показано, що в сильно нерівноважних умовах, т. Е. При досить сильних потоках речовини і енергії, системи можуть мимовільно і стійко розвиватися, диференціюватися. В таких умовах можливі й обов'язкові порушення однозначних причинно-наслідкових зв'язків і прояви ембріональної регуляції та інших явищ. Прикладами дисипативних небиологічних систем є хімічна реакція Бєлоусова - Жаботинського, а також математична модель абстрактного фізико-хімічного процесу, запропонована англійським математиком А. Тьюрингом.
На шляху моделювання морфогенезу як самоорганізованого процесу зроблені перші кроки, а всі перераховані концепції цілісності розвитку носять поки фрагментарний характер, висвітлюючи то одну, то іншу сторону.