| Головна |
| «« | ЗМІСТ | »» |
|---|
З часу перевідкриття законів Менделя генетика неодноразово стикалася з їх «порушеннями», винятками: появою різних результатів в реципрокних схрещуваннях, розщепленням в першому гібридному поколінні, порушенням вільного комбінування генів.
Навіть правило чистоти гамет, яке У. Бетсон сформулював як основу уявлень про генетичну дискретності, не відповідало законам Менделя. Проте аналіз всіх цих винятків служив розвитку ядерної, а потім хромосомної теорії спадковості. Численні експерименти доводили важливу роль ядра і хромосом в спадковості і, здавалося, не залишали місця для інших детермінант спадкових ознак. Проте уявлення про генах поза хромосом в кінці кінців отримали фактичне обгрунтування і розвинулися в самостійний розділ генетики, який досліджує нехромосомной спадкування і так зване цитоплазматичне успадкування.
Любителі кімнатного квітництва добре знають декоративні ряболисті форми аукуби, герані, плюща, хлорофітума, традесканції і інших рослин. У них зелене листя поцятковані білими або жовтими плямами, смугами - ділянками тканин, що не містять пластид або мають дефектні пластиди, позбавлені хлорофілу. Такі ж форми зустрічаються в природі і у культурних рослин: лев'ячого зіва, нічної красуні, примули, кукурудзи та ін.
Спадкування ряболиста на початку XX ст. вивчали К. Корренс (1908) у нічний красуні (Mirabilis jalapa) і Е. Баур (1909) у герані (Pelargonium zonale). Найбільш характерний приклад - успадкування ряболиста у нічної красуні. Ряболисті форми цієї рослини утворюють цілі пагони, позбавлені хлорофілу.
Якщо в якості материнської форми взяти квітки бесхлорофилльной втечі і опиліть їх пилком зеленого рослини, то в F з'являться тільки бесхлорофілльние форми, які незабаром гинуть, т. к. не здатні до фотосинтезу. При реципрокні схрещування в F, всі рослини виявляються нормальними - зеленими. При запиленні квіток ряболисті втечі пилком зеленої форми в Fi утворюються бесхлорофілльние, ряболисті і зелені рослини, при реципрокні схрещування - тільки зелені. Це приклад материнського типу успадкування. Для нього характерне контрастне відмінність між результатами реципрокних схрещувань.
Подібні відмінності в реципрокних комбінаціях спостерігаються при схрещуванні зелених і ряболисті форм і у інших рослин, наприклад у зніту (Epiloblum). В останньому випадку якщо материнська форма зелена, а батьківська пестролистная, то в F, переважна більшість гібридів зелені, але зрідка (з частотою близько 1 на 1 000) зустрічаються і ряболисті форми.
Бувають і інші відмінності при реципрокних схрещуваннях, наприклад у герані: якщо квіти ряболисті рослини запилюють пилком зеленого, то до 30% гібридів ряболисті, 70% - зелені. При реципрокні схрещування 70% гібридів виявляються ряболисті, 30% - зеленими. Це приклад успадкування за батьківським типу.
Пластида - самовідтворюються органели клітини. На відміну від хромосом ядра при розподілі між дочірніми клітинами вони не підкоряються строгим законам мітозу і мейозу. Апарат, керуючий розподілом пластид, в даний час невідомий, і вважається, що вони потрапляють в дочірні клітини випадково при розподілі цитоплазми, завдяки тому, що містяться в клітці у безлічі екземплярів (до декількох сотень).
Реципрокні схрещування рослин розрізняються кількістю цитоплазми, що привноситься в зиготу яйцеклітиною і спермием. При цьому пластиди передаються тільки від материнської форми, як в разі нічної красуні, або зрідка від батьківської форми, як у зніту, можуть передаватися від обох батьків або, нарешті, переважно від батьківської форми, як у герані. Цим і пояснюються відмінності результатів при реципрокних схрещуваннях.
Якщо в цитоплазмі для ряболисті рослин є нормальні хлоропласти, що містять хлорофіл, і дефектні - позбавлені хлорофілу, то при мітозі в деякі клітини можуть потрапити тільки нормальні, а в деякі - тільки дефектні пластиди. Велика частина клітин отримує обидва типи пластид. Цим пояснюються поява забарвлених і нефарбованих ділянок тканини у ряболисті рослин і складні картини розщеплення, іноді спостерігаються при реципрокних схрещуваннях.
У хлоропластах вищих рослин і водоростей виявлена ДНК. З клітин фотосинтезуючої найпростішої водорості Euglena gracilis виділена кільцева молекула хлоропластної ДНК, яка, судячи з її довжині, має молекулярну масу 8,3 х 107 Д (близько 126 т. П. Н.). Щільність цієї ДНК, що дорівнює 1,685 г / см3, відрізняється від щільності ядерної ДНК (1,707 г / см3), Завдяки чому її можна ідентифікувати як окремого піка при рівноважному центрифугуванні в градієнті щільності. Цей пік відсутній у мутанта евглени, що втратив хлоропласт. Порівняльна характеристика щільності ядерної та хлоропластної ДНК різних організмів представлена в табл. 8.1.
Таблиця 8.1
ДНК хлоропластів водоростей і вищих рослин
|
організм |
Щільність ДНК, r / CMJ |
|
|
ядро |
хлоропласт |
|
|
Chlamydomonas |
1,724 |
1,695 |
|
Chlorella |
1,716-1,724 |
1,692-1,695 |
|
Euglena |
1,707 |
1,685 |
|
Porphyra tenera |
1,720 |
1,696 |
|
Nicotiana tabacum |
1,690-1,698 |
1,697-1,698 |
|
Spinacia oleracea |
1,694-1,695 |
1,696 |
|
Brassica rapa |
1,692 |
1,695 |
|
Allium cepa |
1,689-1,691 |
1,696 |
|
Triticum aestivum |
1,702 |
1,698 |
|
Lathyrus odoratus |
1,695 |
1,697 |
|
Lactuca sativa |
1,694 |
1,697 |
Примітка: Цифри через тире вказують на крайні значення, отримані різними авторами.
Виявлення ДНК в хлоропластах стимулювало їх вивчення як напівавтономною самовідтворюється системи. У пластидах виявлений самостійний апарат білкового синтезу, багато в чому відрізняється від цитоплазматичного, характерного для еукаріот, і схожий з апаратом білкового синтезу прокаріотів.
Основні відомості про строгих закономірності пластидної спадковості отримані в експериментах із зеленою одноклітинної водорості СИатукотопаз. У цій водорості в клітці знаходиться один хлоропласт, де виявлені два Фельгена-позитивних тільця. Наявність двох молекул ДНК в хлоропласті (диплоїдний хлоропласта) підтверджують дані генетичного аналізу СИатуАотогш геткагскі. В її пластидної ДНК, або пластому, локалізовані гени, перераховані нижче (табл. 8.2).
Мутації зазначених генів (табл. 8.2) мають наступні фенотипічні прояви:
Таблиця 8.2
мутантні гени СІ1атус1отопаз гапІагЛі, локалізовані в хлоропластної групі зчеплення
|
ген |
фенотип мутанта |
|
ш |
Не зростає при 35 ° С |
|
ас |
Потребує ацетат |
|
ас! |
Потребує ацетат |
|
5 / л4 |
Потребує стрептоміцин |
|
ЗТЗ |
Стійкий до стрептоміцину (500 мкг / мл) |
|
зтя2 |
Стійкий до стрептоміцину (50 мкг / мл) |
|
ПЕА |
Стійкий до неаміціну (1 мкг / мл) |
|
ЄГУ |
Стійкий до еритроміцину (5 мкг / мл) |
|
саг |
Стійкий до карбоміціну (50 мкг / мл) |
|
Стійкий до спіроміціну (100 мкг / мл) |
|
|
С1Е |
Стійкий до клеоціну (50 мкг / мл) |
|
о1е |
Стійкий до олсандоміціну (50 мкг / мл) |
|
БРС |
Стійкий до спектіноміціпу (50 мкг / мл) |
|
сі / |
Умовна потреба в стрептоміцин |
Всі ці мутації зазвичай успадковуються тільки по материнській лінії, т. Е. Від батька тг. Це пов'язано зі складним біогенезом хлоропласта зигот, в результаті якого зберігається лише ДНК хлоропласта, отримана від батька ТЗ. Таке правило має виняток, на якому будується весь рекомбінаційний аналіз хлоропластної групи зчеплення: іноді спонтанно (не частіше ніж 1% випадків) зигота отримує обидва пластому (хлоро пластних генома) - від пи * і ТЗ. Тоді всі чотири (або вісім) зооспори, які утворюються при дозріванні зиготи, виявляються гетерозиготними по генам хлоропласта. Такі гетерозиготи називаються цітогетамі - цитоплазматичними гетерозиготами. Частоту появи цітогет можна підвищити до 50%, опромінюючи жіночі гамети (Пі *) ультрафіолетовим свего безпосередньо перед каплиці.
При утворенні цітогет все хлоропластної маркери, що вводяться в схрещування, успадковуються не по материнській лінії, а від обох батьків, т. Е. т1* і т (. На відміну від ядерних генів, що виявляють мейотічне розщеплення в тетрадах (октадах) 2: 2 (4: 4), хлоропластної гени у цітогет розщеплюються не в мейозі, а при кожному мітотичного ділення зооспор, поки не вийдуть в гомозиготу. Розщеплення відбувається в результаті обмінів на стадії чотирьох ниток, т. Е. В момент, коли молекули хлоропластної ДНК вже подвоєні, але ще не розійшлися в дочірні клітини. При цьому спостерігаються реципрокная рекомбінація, як при митотичному кроссинговере на ділянці ген-центромера, і конверсія. Роль центромери при цьому відіграє точка прикріплення хлоропластної ДНК до мембрани, керуюча розбіжністю ниток ДНК при діленні пластиди. Картування генів у цітогет ведеться трьома способами:
Карта, побудована таким чином, має кільцеву форму (рис. 8.1). Позначення генів представлені в табл. 8.2. Квадратна дужка об'єднує гени, послідовність яких не встановлена.
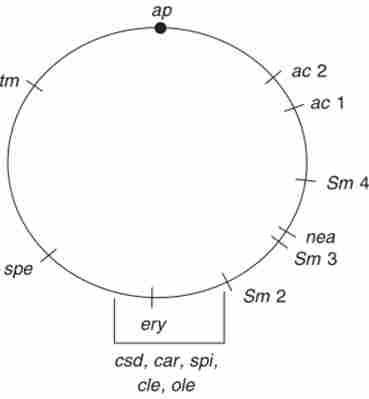
Мал. 8.1. Кільцева група зчеплення хлоропласта СИатус1отопаз геткагЖп: ар - точка прикріплення хлоропластної ДНК до мембрани, ікающая роль центромери
Генетичний аналіз цітогет у хламідомонади заснований не тільки на підвищенні частоти виняткових зигот при опроміненні ультрафіолетовим світлом батька пі але і на селекції виняткових зигот, що дають початок цітогетам. Для цього використовують домінантний хлоропластної маркер батька / я / *. наприклад, т/ * - стрептоміцінзавісімий - ЗС1 (Рецесія), а ТГ - стрептоміцінчувствітельний - 55 (домінант). Зиготи поміщають на середу без стрептоміцину, на якій не можуть проростати регулярні зиготи, що успадковують ознака хлоропласта по материнській лінії (рис. 8.2) - від ТЗ. На цьому середовищі проростають тільки виняткові зиготи, оскільки цітогсти не потребують стрептоміцин.
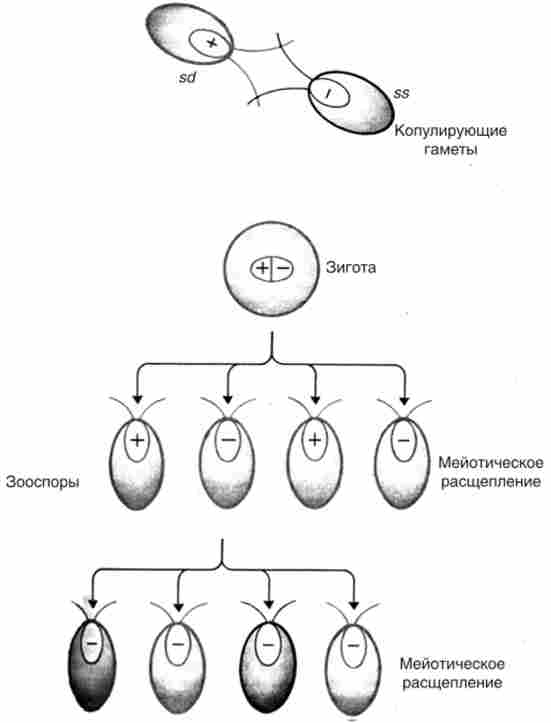
Мал. 8.2. Розщеплення в потомстві винятковою зиготи, що дає цітогети: «+» і «-» - типи спаровування, 55 - чутливість до стрептоміцину,
5 (1 - залежність від стрептоміцину
Такі виняткові зиготи, якщо вони були гетерозиготних по ядерним генам, розщеплюються в мейозі, а гаплоїдні зооспори далі розщеплюються в мітозі по маркерами цітогети.