| Головна |
| «« | ЗМІСТ | »» |
|---|
У багатьох рослин, диких і культурних, зустрічаються форми, що не утворюють пилку або утворюють пилок, нездатну до запліднення. Це явище називається чоловічою стерильністю. Воно може визначатися одним рецесивним геном в хромосомі. Відомі форми чоловічої стерильності, успадковані по материнському типу і отримали назву цитоплазматичної чоловічої стерильності (ЦМС). Материнське успадкування стерильності пилку вперше було виявлено в 1930-х рр. у кукурудзи М. Родсом в США і М. І. Ханджіновим в СРСР.
При запиленні кукурудзи з чоловічою стерильністю пилком нормальних рослин виходило потомство зі стерильною пилком. При повторних зворотних схрещуваннях з рослинами, що мають нормальну пилок, знову виникало потомство з чоловічою стерильністю, навіть якщо все хромосоми материнській лінії заміняли на хромосоми батьківській, нормальної лінії. Таким чином, спадкування за материнському типу і непричетність до цього процесу хромосом дозволили локалізувати в цитоплазмі детермінант, що визначає чоловічу стерильність у кукурудзи.
Завдяки тому що у кукурудзи в основній масі стерильною пилку зустрічаються рідкісні пилкові зерна, здатні до нормального запилення, виявилося можливим реципрокное схрещування. У першому ж гібридному поколінні були отримані повністю фертильні рослини (рис. 8.3).
У кукурудзи відомо кілька типів цитоплазматичної чоловічої стерильності, наприклад техаський (Г), при якому повністю стерильні пильовики не виступають назовні, і молдавський, ши USDA (5), при якому частина або всі пильовики виступають назовні. Ці два типи стерильності розрізняються також за характером взаємодії з генами - восстановителями фертильності. Зокрема, фертильність у рослин з цитоплазмою техаського типу відновлюють два гена в хромосомі II і ряд генів в хромосомах III, IV, VII і X.
Якщо позначити цитоплазматичний фактор стерильності як Суг ' і нормальну цитоплазму як Сур, а домінантну ядерну аллель - відновник фертильності - як /? / (рецесія - rf), то ознака цитоплазматичної чоловічої стерильності розвинеться тільки у рослин rfrfCyf, в той час як RfRfCy?, RfrfCyf, RfRfCyf, RfrfCyt4, rfrfCyf будуть фертильними.
Це явище - відновлення фертильності пилку - широко використовується на практиці для отримання гетерозисних подвійних міжлінійних гібридів кукурудзи (рис. 8.4). Для цього поруч висівають по дві лінії кукурудзи зі стерильною і фертильной пилком. Це забезпечує тільки перехресне запилення, що дуже істотно, оскільки кукурудза само- сумісна при запиленні. Лінії по генам Rf підбирають таким чином, що при одному схрещуванні гібриди мають стерильну, а при іншому - фертильних пилок. При висіві цих гібридів на наступний рік таким же чином отримують подвійні гібриди. При цьому в половині випадків запилення дає фертильні за пилку рослини, як і слід при анализирующем схрещуванні (рис. 8.4).
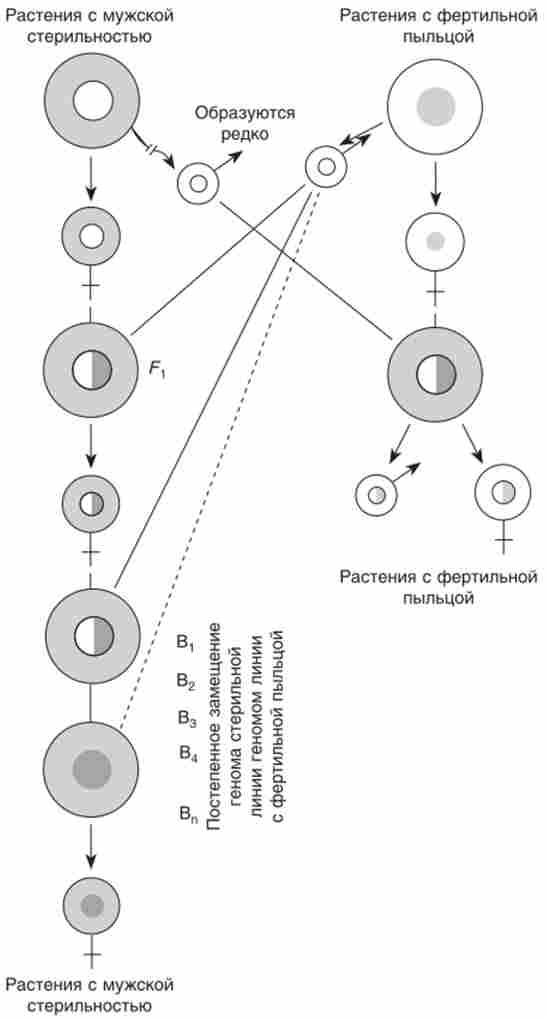
Мал. 8.3. Спадкування чоловічої стерильності у кукурудзи по материнській лінії
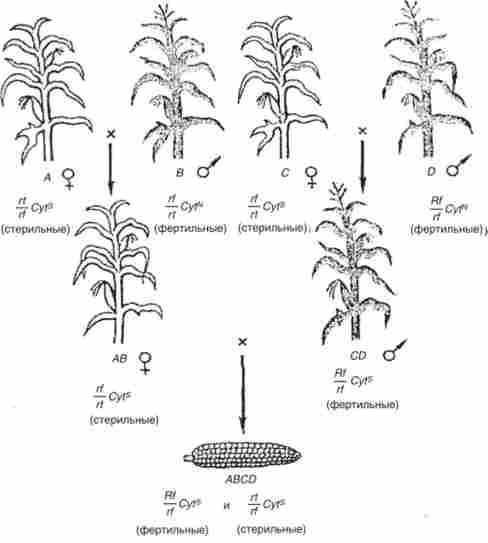
Мал. 8.4. Отримання подвійних гібридів у кукурудзи
Цей прийом економічно дуже вигідний, оскільки дозволяє уникнути кастрації - обламування волоті у кукурудзи, що вимагає великих витрат праці. Широке поширення техаського типу стерильності мало і негативні наслідки, оскільки рослини з такою цитоплазмой виявилися сприйнятливими до грибкових захворювань - гельмінтоспо- ріозу листя, збудником якого є пліснявий гриб Не'гптШропіт таусНь, знищив в 1970 р більше половини врожаю кукурудзи в південних районах США. Токсин, що виділяється цим цвілевих грибів, руйнує внутрішні мембрани мітохондрій у ліній кукурудзи з техаським типом чоловічої стерильності. Це змусило шукати інші типи ЦМС у кукурудзи, щоб використовувати їх в селекції замість техаського типу.
Зв'язок чутливості до гельмінтоспоріозу з мітохондріями, мабуть, виявилася невипадковою. Мітохондрії, як і хлоропласти, мають власну ДНК. В мітохондріях Су? відсутня ділянку довжиною близько 10 000 п. н., присутній в мітохондріях Су?1. Цей фрагмент виявляє гомологію з ДНК хлоропласта.