| Головна |
| «« | ЗМІСТ | »» |
|---|
мітохондрії - це, подібно хлоропластам, самовоснроізводящіеся напівавтономні органели клітини, що містять кільцеві молекули ДНК з різною контурної довжиною. Вони забезпечують дихання клітин рослин, тварин і еукаріотичних мікроорганізмів. ДНК мітохондрій по нуклеотидному складу і внаслідок цього по щільності відрізняється від ДНК ядра (табл. 8.3). Мітохондрії мають власний апарат білкового синтезу, що відрізняється від цитоплазматичного і близьке до апарату білкового синтезу прокаріотів.
Таблиця 8.3
ДНК мітохондрій мікроорганізмів, рослинних і тваринних клітин
|
організм |
Щільність ДНК, г / см! |
|
|
ядро |
мітохондрія |
|
|
дріжджі - Sacch. cerevisiae |
1,698 |
1,684 |
|
найпростіші: |
||
|
Tetrahymena |
1,685-1,692 |
1,684-1,686 |
|
Leishmania henrietti |
1,721 |
1,699 |
|
Tripanosoma cruzi |
1,710 |
1,699 |
|
слизовик Physarum polycephalum |
1,700 |
1,686 |
|
тварини: |
||
|
Жаба - Rana pipiens |
1,702 |
1,702 |
|
короп - Cyprinus carpo |
1,697 |
1,703 |
|
курячий ембріон |
1,701 |
1,707 |
|
голуб - Calumba livia |
1,700 |
1,707 |
|
качка - Anas domestica |
1,700 |
1,711 |
|
Морська свинка - Cavia porcellus |
1,700 |
1,702 |
|
печінка миші |
1,701 |
1,701 |
|
печінка бика |
1,703 |
1,703 |
|
Людина (лейкемичні лейкоцити) |
1,695 |
1,705 |
|
водорості: |
||
|
Chlorella |
1,717 |
1,712 |
|
Euglena |
1,707 |
1,690 |
|
Вищі рослини: |
||
|
Pena - Brassica rapa |
1,692 |
1,706 |
|
Цибуля- Allium сірка |
1,698 |
1,706 |
Багатоклітинні еукаріоти незручні для вивчення генетики мітохондрій, оскільки їх клітини - облігатні аероби, які не можуть існувати при порушенні основної функції мітохондрій - дихання. У той же час дріжджі-сахароміцети є факультативними аеробами. При придушенні дихання вони можуть існувати за рахунок бродіння, використовуючи для цього глюкозу і деякі інші цукру в якості джерел вуглецю На неферментіруемих джерелах вуглецю наприклад на етанолі, гліцерині, лактату кальцію, за відсутності дихання дріжджі не ростуть.
Перші відомості про ознаки, контрольованих мітохондріями, були отримані у дріжджів Saccharomyces cerevisiae в кінці 1940-х рр. в лабораторії Б. Ефруссі. У цих грибів відомі мутантні форми, що утворюють на глюкозі дрібні колонії, так звані Pe / i'ie-мутанти, фенотип яких позначають Реєстр на відміну від дикого типу Реєстр. мутанти Реєстр не ростуть на неферментіруемих джерелах вуглецю, оскільки не здатні до дихання. Схрещуючи гаплоїдні клітини Реєстр х Реєстр, можна отримати гібриди дикого типу, здатні до дихання. Зошитовий аналіз таких гібридів (рис. 8.5) показує, що ознака Реєстр від незалежно отриманих мутантів успадковується по-різному. Одні гібриди показують нормальне розщеплення (2Реєстр : 2Реєстр), а інші не виявляють розщеплення в тетрадах (4Реєстр : 0Реєстр). Очевидно, в першому випадку нездатність до дихання визначається хромосомної мутацією, а в другому - нехромосом- ної, мабуть, цитоплазматичної. Ці два типи мутантів Реєстр були названі відповідно генеративних і вегетативними.
Нехромосомной природу вегетативних ЛгГ-мутаітов підтвердили і багаторазові зворотні схрещування сегрегантов Реєстр з батьком Реєстр. У всіх випадках ознака Реєстр в тетрадах не проявляє (рис. 8.5, б), в той час як по ядерних маркерами, введеним в схрещування, спостерігали регулярне розщеплення 2: 2. При схрещуванні вегетативних та генеративних мутантів Реєстр утворюються гібриди Реєстр, в тетрадах яких відбувається розщеплення 2Реєстр: 2Реєстр.
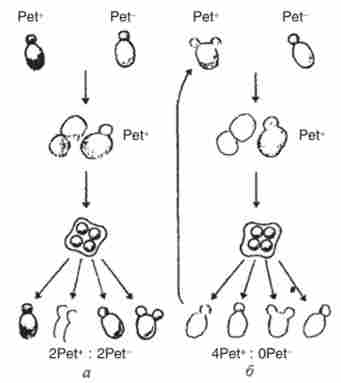
Мал. 8.5. Зошитовий аналіз здатності до дихання у гібридів між гаплоїдії дикого типу: а - генеративних (ядерними); б - вегетативними (неядерними) / 'ЕГ-мутантами
Вегетативні Pef-мутанти виникають спонтанно. Іноді вони складають до 1% культури. Їх поява стимулюють висока температура, акрифлавіну, бромистий етидій в однаковій мірі у гаплоидов і диплоидов. При пересіву ці мутанти ніколи не ревертіруют до фенотипу Pet+ на відміну від генеративних Pei. Зазначені впливу не індукують генеративних мутантів Pei. Все це змусило припустити, що вегетативні Pei - результат втрати якогось детермінанта, що знаходиться в цитоплазмі, можливо, мітохондрії. Відкриття мітохондріальної ДНК (мтДНК) дозволило перевірити це припущення.
Порівняння мтДНК з штамів дикого типу і з вегетативних мутантів Pei показало, що останні несуть делеции мтДНК різної протяжності аж до повної її втрати. Надалі в якості генотипичного символу позначення pet зберегли тільки для рецесивних алелей ядерних генів, яких тепер відомо понад 20. Мітохондріальні мутації стали позначати rh
При генетичному аналізі ознак, контрольованих мтДНК, виявився ряд особливостей поведінки мітохондріальних генів. При спарюванні гаплоїдних клітин, що розрізняються по аллелям будь-якого мітохондріального гена, утворюється популяція диплоидов, що складається з клітин, які отримали той чи інший аллель, причому в співвідношенні, характерному для схрещується штамів. Відсоток диплоидов, які отримали певний аллель, позначають як частоту трансмісії даного аллеля.
При дослідженні рекомбінації мітохондріальних геномів потрібно враховувати, що в зиготі створюється популяція молекул мтДНК, що вступають в багаторазові спарювання і обміни, аналогічно тому, що відомо про рекомбінації ДНК бактеріофагів. При перших діленнях зиготи ця популяція досить швидко розщеплюється, так що диплоїдні вегетативні клітини містять тільки один тип молекул мтДНК: один з батьківських чи рекомбінантний. При цьому для деяких маркерів спостерігається явище полярності рекомбінації, що виражається в тому, що порушується рівність реципрокних рекомбінантних класів.
Явища різної трансмісії і полярності рекомбінації маркерів ускладнюють кількісну оцінку частоти рекомбінації і картування генів. Ефективним способом побудови генетичної карти мітохондрій виявився метод, заснований на використанні г / го-мутацій, що представляють собою делеции. При цьому досліджують частоту спільної втрати або збереження досліджуваних маркерів у незалежно отриманих мутантів rho ~. Таким способом визначають чергування маркерів на карті. Крім того, існують набори штамів дріжджів гЬоГ з фізично охарактеризованими поділами мтДНК. Досліджувану мутацію і делецию об'єднують при схрещуванні і перевіряють, чи виникають з досліджуваного гену рекомбінанти дикого типу в постзіготічних мітозу. Відсутність рекомбинантов означає, що ділення захоплює досліджуваний ген.
Якщо є тестери з перекриваються поділами, захоплюючими як різні, так і однакові ділянки мтДНК, в сумі складають весь мітохондріальний геном, то в якісному тесті можна картировать будь-яку мутацію мтДНК (табл. 8.4). Карта мітохондріального геному має кільцеву форму (рис. 8.6).
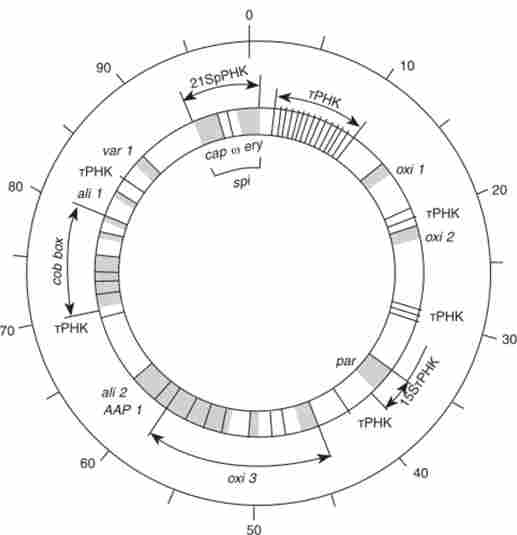
Мал. 8.6. Карта мітохондріальної групи зчеплення 8асск. сегечИае. Цифри зовнішнього кола маркують ділянки загальною шкали в 100 одиниць, яка відповідає 75 000 п. Н.
Виділення і дослідження мтДНК дозволило вирішити ще одну загадку вегетативних Реєстр-мутантів, з якою зіткнулися Б. Ефруссі і його колеги (1955). При схрещуванні клітин деяких г /? про "-мутанта з клітинами дикого типу з різною частотою (до 99%) утворюються диплоидов, не здатні до дихання. Це властивість, назване супресією остю, стабільно наследовалось при вегетативному розмноженні і при схрещуванні.
Таблиця 8.4
Гени, картірованниє в мітохондріальної групі зчеплення у дріжджів
|
ген |
контрольований ознака |
|
СО |
Фактор полярності рекомбінації |
|
215 |
РНК великої субодиниці рибосом |
|
cap |
Стійкість до хлорамфеніколу |
|
spi |
Стійкість до спіроміціну |
|
ery |
Стійкість до еритроміцину |
|
15 S |
РНК малої субодиниці рибосом |
|
tRNA |
тРНК |
|
25 генів: |
|
|
cob |
Апоцітохром Ь |
|
oxil |
субодиниці |
|
axil |
Цитохром-с- |
|
oxB |
оксидази |
|
olil |
Субодиниці АТФази, |
|
oia J |
мутації стійкості до олігоміціну |
|
aap 1 |
субодиниця АТФази |
|
vari |
Білок малої субчастіци рибосом |
У зигот дріжджів можна індукувати мейоз (що досягається простим перенесенням їх на середу з ацетатом калію або натрію) відразу після їх утворення від схрещування клітин супресивний л7го'-мутантів і клітин дикого типу. В цьому випадку мейоз відбувається нормально і можливий зошита аналіз (рис. 8.7), хоча після вегетативного брунькування зигот і освіти диплоїдних клітин останні в 99% випадків мають фенотип Pei і вже не здатні до мейозу і спорообразованію.
Зошитовий аналіз Супресивна показує відсутність розщеплення в переважній більшості тетрад (0Pei *: 4. Реєстр). Виявилося, що у високосупрессівних мутантів зберігається невелика ділянка мтДНК з точкою початку реплікації. Такі міні-кільця реплицируются швидше нормальної мтДНК і швидко витісняють її при вегетативному діленні клітин. Так звані нейтральні гАо'-мутанти, що не володіють властивістю Супресивна, взагалі не містять мтДНК. Це ДАТ-мутанти.
Генетика мітохондрій найкраще розроблена для дрожжей- сахароміцетів, однак ряд прикладів мітохондріального успадкування отримано і у інших об'єктів.
У Neurospora crassa мутація року (Убогий, кволий) призводить до зміни морфології мітохондрій, порушення в них білкового синтезу, відсутності деяких мітохондріальних ферментів. Зовні це виражається в повільному зростанні штаму. Ознака успадковується по материнській лінії. Це можна встановити завдяки доступності реципрокних схрещувань у нейроспори. Роль чоловічих гамет, практично не вносять цитоплазму при заплідненні, грають мікроконідії. мутація року", Мабуть, результат зміни мітохондріального геному.
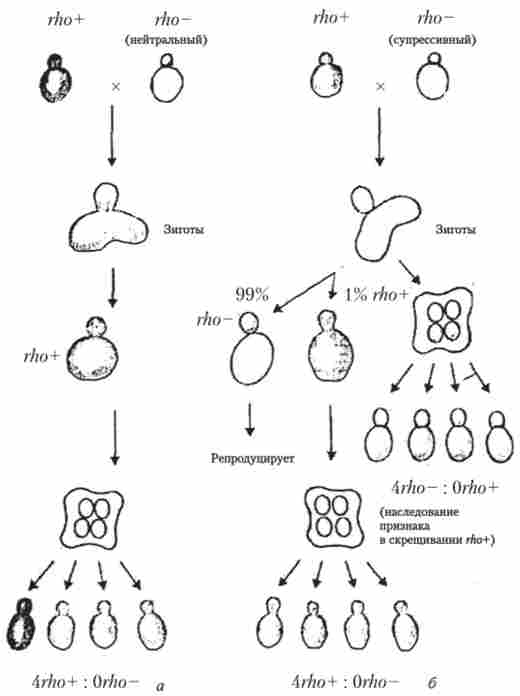
Мал. 8.7. спадкування ознаки Pet ' у дріжджів Saccli. cerevisiae в скрешіваніі rho ~ х rho *: а - нейтральний; б - супресивний
У іншого аськоміцетов - Podospora атеппа (життєвий цикл Р. атеппа і N. сгм5а подібний) - добре відомий ознака «Старіння міцелію», т. е. поступове зниження життєздатності при безперервному вирощуванні. Тривалість життя різних штамів варіює від 9 до 106 днів.
Старіння успадковується по материнській лінії. Його визначає інфекційне початок, як показали досліди по зараженню нормального міцелію в результаті його злиття з старіючої гифой (рис. 8.8). Показаний фенотип культур, отриманих з фрагментів: а - відсутність зростання, Ь - зростання тільки в мікрокраплі, з - зростання менше 2 см, а? - зростання до 2-4 см, в - зростання до 4-6 см, / - зростання до 6-8 см, g- зростання до 10-20 см, А - нормальна тривалість життя.
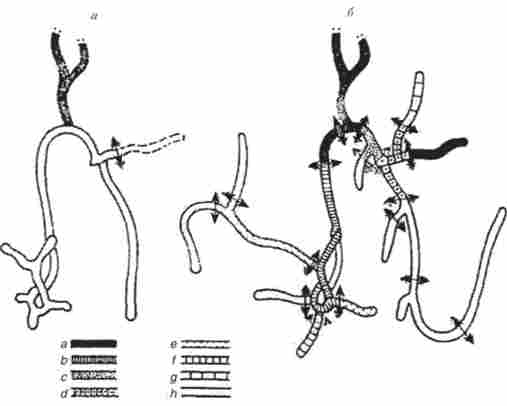
Мал. 8.8. Вивчення зараження нормального міцелію при злитті його з старіючої гифой (чорна) Ройозрога апзеппа: а - стан міцелію безпосередньо після злиття; б-стан міцелію через 12 год, коли його розділили на фрагменти (показано стрілками)
Прискорене старіння спостерігали в ділянках міцелію, безпосередньо прилеглих до точки анастомозу. У міру віддалення від неї вираженість ознаки слабшала. старіння гіф Р. апеегта обумовлено поширенням плазміди, инфицирующей мітохондрії.