| Головна |
| «« | ЗМІСТ | »» |
|---|
Існування напівавтономних клітинних органел зі своїми генетичними детермінантами, а також присутність в клітці інших генетичних детермінант, локалізованих поза ядра, спонукає звернутися ще до однієї сторони парасексуальними процесу. В ході цього процесу, ініційованого злиттям гіф і освітою гетерокаріонов, можуть об'єднуватися різні по генетичній характеристиці неядерні елементи. Вони по-різному комбінуються один з одним і з ядрами, що відбивається на загальній картині вегетативного розщеплення. Приклади такої перекомбі- нації ядер і неядерних елементів отримані у міцеліальних грибів Neurospora, Aspergillus, Podospora та ін. Однак саме природний стан многоядерного міцелію у цих об'єктів створює труднощі у вивченні процесу такого вегетативного розщеплення.
Більш суворий підхід до вивчення перекомбінації ядер і неядерних елементів розроблений для дріжджів-сахароміцетів, у яких при гібридизації також утворюється, хоча і короткочасна, стадія гетерокаріонов. Р. Райт і Дж. Ледербергом (1957) використовували це явище для вивчення рекомбінації неядерних детермінант ознаки Pet і ядерних маркерів. Вони показали, що перші бруньки зиготи дріжджів, отримані від схрещування клітин rho ~ х г ho *, іноді можуть виявитися гаплоїдними і нести ядра того чи іншого з батьків, судячи з рецесивним маркерами, в цитоплазмі rho *. Це явище освіти ядерно-цитоплазматичних гібридів І. А. Захаров (1969) назвав цітодукціей.
Цітодукція виявилася зручною для локалізації генів в ядрі або цитоплазмі, особливо в тих випадках, коли вони виявляють неменделевская спадкування в мейозі. Цітодукція зазвичай відбувається з частотою менше 1%, т. Е. Менше 1% всіх утворюються зигот відгалужується цітодуктанти - гаплоїдні клітини зі змішаною цитоплазмою. В експериментах по цито- продукції використовують клітини одного з батьків, марковані з мітохондрій нейтральної (НЕ супрессивной) мутацією rho0. Цей же штам зазвичай несе будь-яку рецесивну ядерну мутацію стійкості, наприклад до канаваніну (сап Г) - аналогу амінокислоти аргініну, або інший рецесивний маркер, за яким легко вести селекцію. Ці клітини - реципієнти цитоплазми. Другий з батьків має нормальні мітохондрії (Rho *) і несе ядерний домінантний аллель чутливості до того ж агенту (CAN Is). Ці клітини - донори цитоплазми. Крім того, ядра обох гаплоидов маркують декількома неселективними рецесивними мутаціями, щоб можна було стежити за поведінкою хромосом при подальшому аналізі.
Цітодуктанти відбирають на середовищі з етанолом (або іншим нефермен- тіруемим джерелом вуглецю) і канаваніном при схрещуванні (рис. 8.9) донора і реципієнта цитоплазми. На цьому середовищі не можуть рости ні донор (він чутливий до канаваніну), ні реципієнт (rho0) цитоплазми, ні нормальний гібрид, який теж чутливий до канаваніну (CAN 1 s/ can l '). Виростають тільки гаплоїдні цітодуктанти, що зберігають рецесивну стійкість і набувають нормальні мітохондрії (Rho *).
Для підвищення частоти цітодукціі використовують ядерні мутації, що затримують каріогамія. Наприклад, ядерна мутація каг 1 підвищує частоту цітодукціі до 80% і більше.
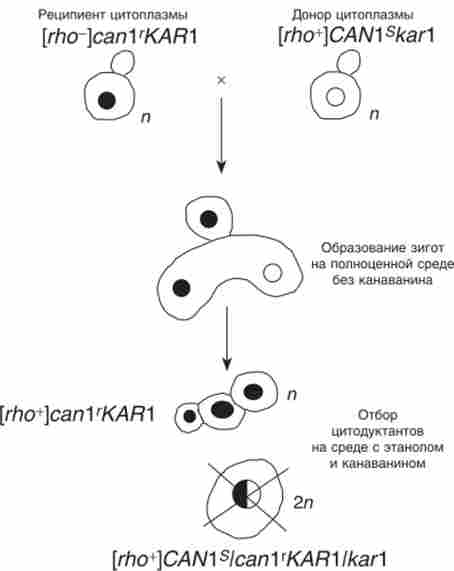
Мал. 8.9. Освіта і селекція цітодуктантов при затримці каріогамії у дріжджів
У дослідах Р. Райта і Дж. Ледербергом за допомогою цітодукціі показана неядерна природа> 7ют-детермінантів. Надалі цей метод був використаний для встановлення ядерної локалізації криптичної (яка не має свого фенотипичного прояви) плазміди з контурної довжиною 2 мкм у дріжджів, а також для встановлення ядерної локалізації сконструйованих на її основі штучних плазмід з ящірний генами.
Ці плазміди при цітодукціі з мітохондріями не передаватимуться або повторно супроводжують ядерні гени того з батьків, з яким вони введені в схрещування. При звичайному гибридологічний аналізі по домінантним плаз- мідно генам спостерігається різке переважання домінантного фенотипу: тетради 4А : 0<я или ЗА: 1а. Це пояснюється великим числом копій плазміди в ядрі і, відповідно, великим числом копій домінантного алеля.
Дана обставина слід підкреслити особливо, т. К. Воно показує, що порушення типового хромосомного спадкування ще не доводить неядерну природу досліджуваного гена.
Застосовуючи цітодукцію, доводять цитоплазматичну локалізацію двунітевой РНК - вірусоподібні детермінанта вбиває активності, виявленого у деяких штамів дріжджів-сахароміцетів. Такі дріжджі при спільному вирощуванні вбивають клітини чутливих штамів.
Генетичний аналіз, заснований на цітодукціі, широко використовується при дослідженні дріжджів і може бути поширений на інші об'єкти.