| Головна |
| «« | ЗМІСТ | »» |
|---|
Гетерозис - гібридна потужність, що виявляється в перевазі гібрида над обома батьківськими формами. Це явище було описано І. Г. Кельрейтер, одним з перших попередників Г. Менделя. Свої результати по вивченню гібридів між «Віргінські» і «перувианского» сортами тютюну він опублікував в 1772 р
Гетерозис широко використовується в селекції рослин і тварин, але механізм гібридної потужності до сих пір до кінця не з'ясований.
Депресія, пов'язана з інбридингом, - явище, протилежне гетерозису. Існування першого зазвичай служить гарантією другого, і якщо інбредних депресії пов'язують з гомозиготизації ліній, то гетерозис - з різким підвищенням гетерозиготности. І. Г. Кельрейтер відзначав, що потужність гібридів пов'язана зі ступенем генетичного відмінності їх батьків і має особливе значення в природних, природних умовах. Ці спостереження в подальшому отримали численні підтвердження. Значення перехресного запліднення детально обговорював Ч. Дарвін (1876) в своїй праці «Дія перехресного запилення і самозапилення в рослинному світі», де зазначав, що перехресне запліднення зазвичай корисно, а самооплодотворение шкідливо. При цьому Ч. Дарвін не залишив без уваги і самозапильні рослини, благополучно існують в умовах інбридингу.
Все це підкреслює складність проблеми гетерозису Характерна його риса - поступове згасання в ряду поколінь (рис. 14.5). За даними
В. Шелла, врожайність зерна гетерозисних гібридів кукурудзи bF2b середньому знижується на 35%, а в F} - на 50% в порівнянні з урожайністю в F. Численні спроби «закріплення» гетерозису поки не привели до істотних результатів, за винятком тих випадків, коли матеріал можна розмножувати вегетативно. Шляхи закріплення гетерозису бачать в перекладі гібридів до розмноження за допомогою апоміксиса або поліплоіді- зації з використанням колхіцину або деяких мутантів з порушеннями мейозу, наприклад мутанта кукурудзи типу elongate, у якого утворюються нередуцірованних (диплоїдні) яйцеклітини. Очевидно, поліплоіді- зация не закріплює гетерозис, а уповільнює зниження його ефекту в наступних поколіннях завдяки порушенню принципу чистоти гамет.
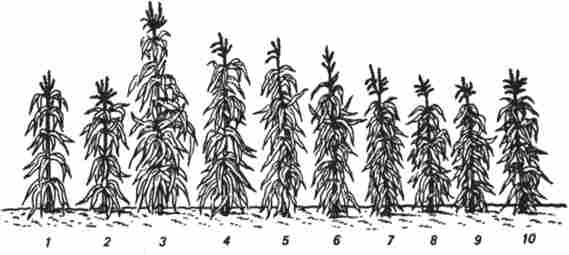
Рис 14.5. Прояв гетерозису в різних поколіннях гібридної кукурудзи: 1-2 - вихідні батьківські форми, 3 - гібридні / ц,
4-10 - гібриди наступних поколінь
Для використання явища гібридної потужності доводиться знову отримувати гетерозисних гібриди.
Гетерозис може стосуватися аж ніяк не всіх ознак рослини або тварини. А. Густафссон запропонував таку класифікацію типів гетерозису у рослин:
Прояв гетерозису залежить від напрямку схрещування і може спостерігатися тільки в одному з реципрокних схрещувань. Велике значення мають також умови вирощування гібридів /г]. Ознаки високої продуктивності можуть в повній мірі проявитися тільки при сприятливих умовах.
Найкращі результати дає гетерозис при схрещуванні певних ліній, тому необхідно попередньо перевіряти їх на комбінаційну здатність, т. е. на здатність утворювати продуктивні гібриди.
Теорії гетерозису поділяють на такі основні групи.
теорія домінування пояснює гетерозис підбором у гібрида сприятливих домінантних алелів різних генів, втрачених при інбридингу. Якщо схрещувати лінії гомозиготні по рецесивним аллелям різних генів, то гібриди виявляться полігетерозіготамі, в яких домінантні атлелі будуть взаємодіяти з комплементарному типу. Якщо врахувати, що кількісні ознаки успадковуються полигенно, то підбір більшого числа адитивно діючих домінантних генів повинен привести до більш потужному їх прояву. Ця теорія була запропонована в 1908 р Г. Девенпорт і розвинена в 1917 р Д. Джойсом. Подальший розвиток популяційної генетики, особливо встановлення гетерозі- готності особин з природних популяцій з багатьох рецесивних аллелям, надають в гомозигот негативний вплив на життєздатність, змусило переглянути цю теорію.
теорія сверхдоминирования пов'язує гібридну потужність з перевагою гетерозиготного стану (АЛ<Лааа). При цьому ефект сверхдоминирования в гегерозіготе може спостерігатися навіть у тому випадку, коли рецесивний аллель в гомозигот летален або призводить до зниження життєздатності. Перевага гетерозиготности добре демонструє приклад підвищення стійкості гібридів льону до іржі. Ця ознака контролюють кілька генів, кожен з серією алелей. При цьому в конкретному гені різні аллели визначають стійкість до різних рас паразита. Гетерозигота по гену стійкості (М'М7) виявляється несприйнятливою до обох рас іржі, імунітет до яких визначають аллели, що знаходяться в гетерозиготі.
У розглянутому прикладі очевидна перевага гетерозиготности, обумовлене незалежним дією алелей. Відомо, що аллели щонайменше половини вивчених генів можуть взаємодіяти по типу межаллельной комплементации. Освіта гібридних білків-мультімеров в природних умовах характерно для великого числа ізозімов. У гібридів описано підвищення активності ферментів і розширення їх ізозімних спектрів. Так, у гібридів кукурудзи виявлено підвищення активності і стабільності алкогольдегідрогенази, а у дрозофіли - окта- ноддегідрогенази в порівнянні з аналогічними показниками схрещується ліній.
Значення алельних взаємодій в прояві гібридної потужності підтверджують приклади так званого моногенного гетерозису. Так, С. Даскалов отримав гетерозисних гібриди томата, перевага яких по продуктивності було пов'язано з гетерозиготністю по одній хлорофільних мутації Хатіаі (Летальної в гомозигот).
Синтез уявлень про алельних і неалельних взаємодіях, що забезпечують гетерозис, запропонував В. А. Струнников в своїй теорії компенсаторних комплексів. Цей термін означає комплекс генів і їх алелів, який підбирається при отриманні інбредних ліній. Компенсаторний комплекс генів нівелює негативні ефекти високого рівня гомозіготносгі, а при гібридизації дає той ефект в вираженні ознак, який і зумовлює гетерозис. Істотно доповнює теорію гетерозису В. А. Струнникова його гіпотеза про стабілізацію дії генів у гібридів в умовах гетерогенної навколишнього середовища і про зменшення впливу спонтанних чинників модифікації.