| Головна |
| «« | ЗМІСТ | »» |
|---|
Всі ферменти біосинтезу жирних кислот утворюють єдиний агрегат - Мультиферментний комплекс, який отримав назву синтетази жирних кислот. Комплекс синтетази тваринних тканин і дріжджів - це димер, що складається з двох ідентичних мономерів, кожен з яких представляє поліпсптідную ланцюг, що включає шість активних центрів ферментів і ацил-переносить білок (HS-АПБ). Ці ферменти утворюють настільки компактну структуру, що вони не піддаються фракціонування і всі спроби виділити їх в індивідуальному вигляді не увінчалися успіхом.
Інший тип структурної організації ферментів синтезу жирних кислот зустрічається у мікроорганізмів і рослин, з яких окремі ферменти можуть бути виділені методами білкового фракціонування.
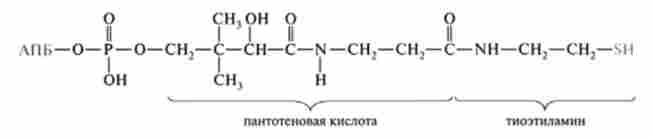
До складу синтетази входить ацил-переносить білок (HS-АПБ). Він має молекулярну масу близько 10 kDa; порівняно термостабилен; містить фосфорильовану пан- тотеновую кислоту (вітамін В3) І тіоетіламін, ковалентно приєднані до поліпептидного ланцюга АП Б через серинові залишок фосфоефірную зв'язком. Слід зазначити, що та ж сама угруповання входить до складу HS-KoA:
Реакційної групою АП Б є HS-група, тому його прийнято позначати у вигляді HS-АПБ. У синтетазної системі цей білок виконує функцію, подібну до HS-KoA, т. Е. Бере участь в передачі ацильних радикалів від одного ферменту до іншого.
Ферменти синтетазної комплексу (в дужках позначені номери каталізуються ними реакцій) наведені нижче:
В процесі біосинтезу жирних кислот для прояву синтетазної активності необхідна участь двох сульфгідрильних груп комплексу. Одна реакційна HS-група належить HS-АПБ одного мономера, інша - залишку цистеїну, що входить до складу р-кетоацил-АПБ-синтази іншого мономера, які розташовані в безпосередній близькості один від одного. У зв'язку з цим синтетазної комплекс активний лише у вигляді димеру. Оскільки кожен з мономерів включає всі ферменти синтетазної комплексу, одночасно на димере йде синтез двох молекул жирних кислот. У наведених нижче реакціях цей комплекс схематично може бути зображений таким чином:

Центральну роль в синтезі жирних кислот в цитозолі за участю ферментів сінтстазного комплексу відіграє синтез пальмітинової кислоти СН3(СН2)] 4СООН, оскільки подальше нарощування ланцюга насиченою жирної кислоти відбувається в мітохондріях шляхом звернення реакцій (3-окислення.
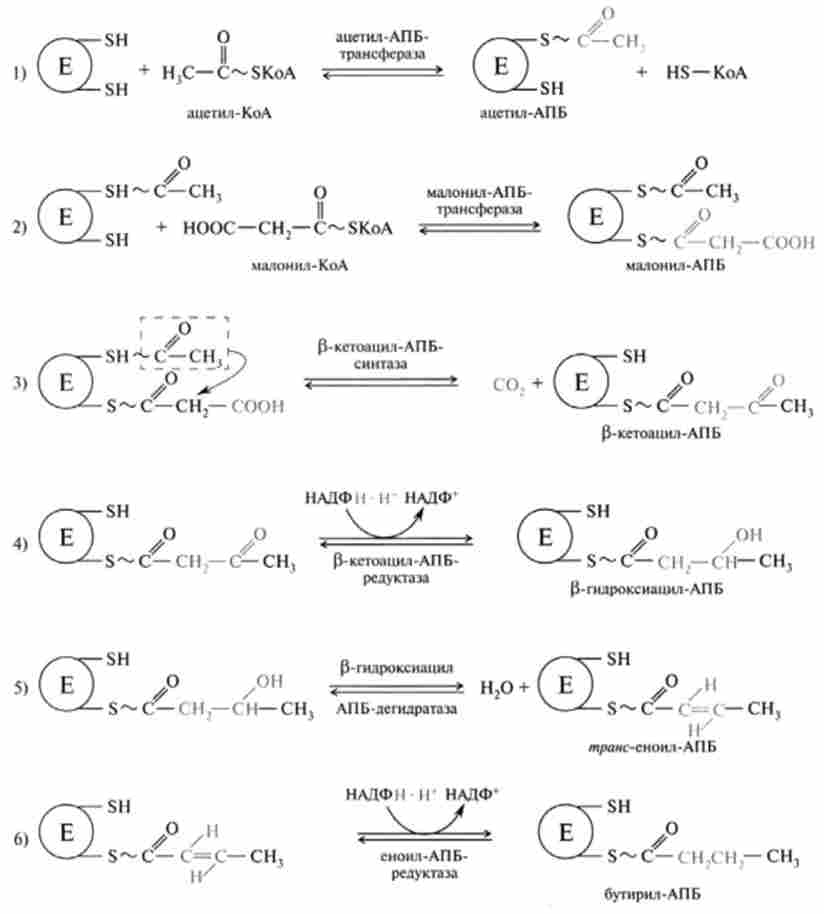
Мал. 23.15. Послідовність реакцій синтезу бутіріл-АПБ
В реакції 1 (рис. 23.15) йде перенесення ацетилу на сульфгідрильні групи цистеїну, Малоні ж взаємодіє з сусідньої HS-групою АП Б, локалізованого на іншому мономере (реакція 2). Центральної реакцією є реакція конденсації (3) ацетил з метиленової групою Малоні, що супроводжується відщепленням С02. В результаті відбувається утворення четирехуглеродного ацильного залишку і вивільнення HS-групи цистеїну, раніше зайнятої ацетильной групою. Декарбоксилирование в цій реакції має енергетичне значення, дозволяє реакції пройти до кінця і, таким чином, є рушійною силою біосинтезу. В подальшому йде відновлення р-кетоацільной групи (реакція 4), потім дегідратація (реакція 5) і знову відновлення ненасиченого / я /? л // з-еноіл-АПБ, в результаті чого утворюється насичений ацил-АПБ (реакція 6). Три останні реакції подібні з відповідними зворотними реакціями р-окислення, відрізняються утворенням в четвертій реакції D-ізомери p-гідроксикислоти, а не L-ізомери, а також тим, що відновником є НАДФН, а не НАДН. Завершується перший етап синтезу переміщенням насиченого ацильного радикала на вільну HS-групу цистеїну, а нова молекула Малоні-КоА взаємодіє з HS-АПБ, цикл реакцій повторюється ще шість раз, і кожен новий залишок малоната, декарбоксіліруясь, вбудовується в вуглецевий ланцюг до тих пір, поки не утвориться 16-вуглецевий пальмитоил-АПБ (рис. 23.16). Завершується синтез кислоти гидролитичним отщеплением HS-АПБ від пальмитоил-АПБ при дії ферменту деацілази, що не входить до складу комплексу синтази жирної кислоти.