| Головна |
| «« | ЗМІСТ |
|---|
В даний час найпростіші (Protozoa) виділяють в самостійне підцарство, що включає 7 типів і налічує понад 65 000 видів, з них понад 10 000 видів становлять паразитичні прості. Генетика їх майже не розроблена.
Характерна риса найпростіших - ядерний дуалізм, т. е. співіснування в одній клітці - організмі так званого генеративного ядра - диплоидного микронуклеуса (Ми) і у високому ступені поліплоїдного (до декількох сотень п) вегетативного ядра - макронуклеуса (Ма). Поліплоїдний вегетативне ядро функціонує в ході вегетативного розмноження клону. Високий рівень плоїдності ма перешкоджає виділенню і дослідженню рецесивних алелей. Генеративное ядро - сховище генетичної інформації - здійснює свої функції під час статевого процесу - кон'югації.
Система статевих типів може бути біполярної, як, наприклад, у Paramecium aurelia, у якій протилежні статеві типи успадковуються як рецесивний (Рр) і домінантний ознаки (Яг, RR), контрольовані алелями одного гена. Відомі випадки множинних статевих типів: наприклад, у Р. bursaria зустрічаються тстраполярная система, контрольована двома генами, і система з восьми статевих типів, контрольована трьома генами. В останньому випадку різниця хоча б по одному з трьох генів визначає приналежність до окремого статевою типу. У іншого роду інфузорій Euplotes відома серія множинних алелів локусу типів спарювання (mt), В якій приналежність до статевого типу визначається домінантним алелем в ряду: mt5 mtA т?> mi1 > mt де знак> означає домінування.
Кон'югацію між клітинами різних статевих типів розглянемо на прикладі Paramecium caudatum. У неї вегетативні клітини об'єднуються в пари (рис. 7.14). Мікронуклеус зазнає два ділення мейозу. Три утворилися гаплоїдних ядра дегенерують, а одне ще раз ділиться мітотично. Одне з утворених ядер мігрує в клітку партнера. Цей процес відбувається реципрокно. Надійшло ядро зливається з власним ядром клітини-реципієнта, утворюючи синкарион. Після обміну про нуклеусами клітини розходяться. Після цього синкарион ділиться кілька разів мітотично, і утворилися ядра диференціюються на ма і Мі. При цьому в ма відбувається ендомітотічна полиплоидизация. Старий макронуклеус дегенерує.
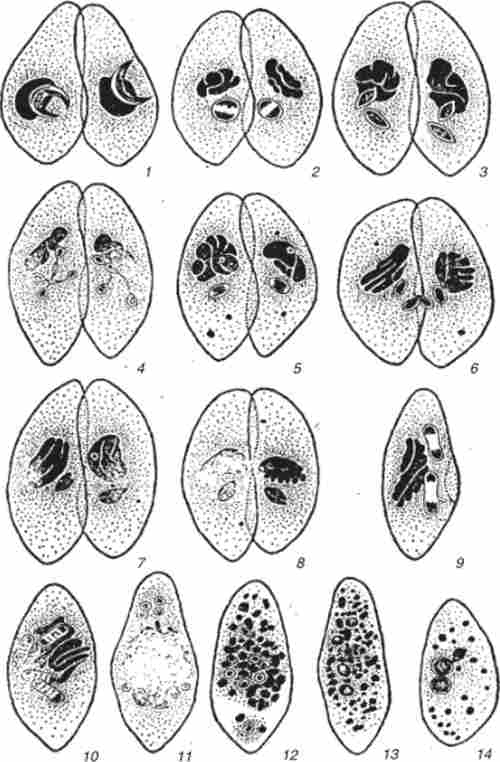
Мал. 7.14. кон'югація Paramecium caudatum: 7,2 перший розподіл мейозу ми; 3,4 другий розподіл мейозу Л / м; 5 - метафаза мітотичного поділу Mir, б - обмін пронуклеуса; 7 - злиття пронуклеусов; 8-10 - розподіл синкариона;
11 - диференціювання дериватів синкариона на чотири зачатка ма в передній частині клітини і на чотири зачатка ми в задній частині клітини; 12 - фрагментація старого ма; 13 - початок розвитку зачатків нового ма; 14 - нащадок першого поділу ексконьюганта з двома зачатками ма і одним ми
Реорганізація ядерного апарату найпростіших може відбуватися не тільки при кон'югації, а й в результаті автогамія (рис. 7.15). Наприклад, у Paramecium aurelia при автогамія реорганізація ядерного апарату відбувається в одній клітці. В результаті мейозу і подальшого мітозу ми утворюються вісім гаплоїдних ядер, з яких два зливаються. Диплоїдний синкарион ділиться і диференціюється на ма і Мі.
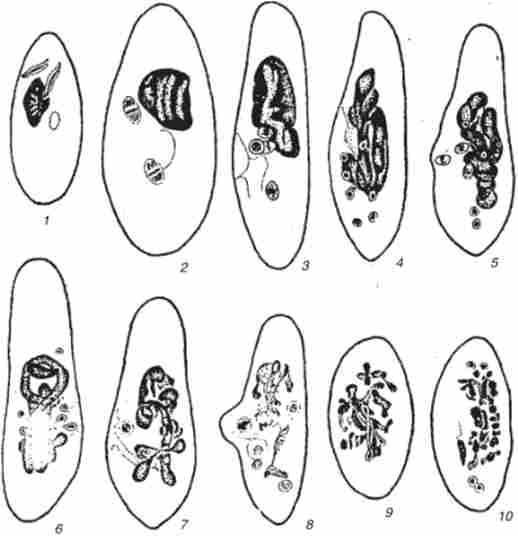
Мал. 7.15. автогамія у Paramecium aure / ia: 1-2- перший розподіл мейозу ми;
3 - другий розподіл мейозу ми; 4 - вісім продукції Ктов другого поділу ми, початок фрагментації ми; 5 - мстафаза третього (мітотичного поділу) ми; б - пронуклеуси в пероральному конусі; 7 - синкарион; 8-9 - перше і друге ділення дериватів синкариона; 10 - чотири дериватів синкариона перед початком диференціювання на ми і ма
Таким чином, кон'югація - це процес, аналогічний перехресному заплідненню, а автогамія - самооплодотворению. Процеси, що ведуть до рекомбінації у найпростіших, мабуть, дуже різноманітні. У деяких з них на відміну від Paramecium спостерігається так звана тотальна кон'югація, у інших відбувається тільки обмін ядрами, у третіх - і обмін цитоплазмой і т. д. Численні варіації в здійсненні мейозу аж до одноступінчастого мейозу у деяких форамініфер, коли число хромосом редукується за одну поділку, без попередньої реплікації.
Існує велика кількість видів Protozoa, позбавлених статевого процесу. До їх числа відносяться і різні види амеб. Для генетичного аналізу у агамное Amoeba proteus А. Л. Юдін запропонував метод мікрохірургічної пересадки ядер (рис. 7.16), що дозволяє досліджувати ядерно цитоплазматичні відносини і їх генетичні наслідки. Цим методом було виявлено ядерно-цитоплазматична несумісність між різними клонами одного виду, а також доведена ядерна детермінація цілого ряду спадкових ознак у A. proteus: Стійкість до метионину, етилового спирту, форма клітини, теплостійкість, швидкість прикріплення до субстрату.
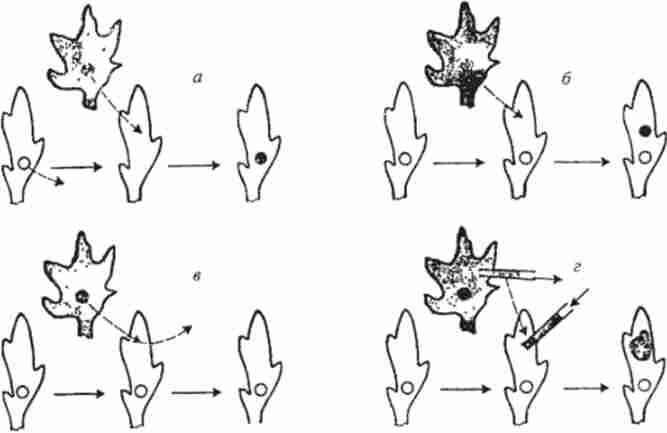
Мал. 7.16. Основні типи мікрохірургічних операцій, що застосовуються при вивченні ядсрних і ядсрно-цітоплазматічсскіх відносин у амеб: а - отримання ядерно-цітоплазматмчного «гібрида»; б - отримання гетерокаріонов; в - тимчасова імплантація ядра; г - трансплантація цитоплазми
Л. Н. Серавін і А. В. Гудков описали у деяких видів амеб явище паракопуляціі, яке полягає в об'єднанні двох багатоядерних особин, потім в їх повне злиття і через деякий час поділі на дві. Генетичні наслідки такої паракопуляціі ще не вивчені.
Розглянувши деякі найбільш вивчені процеси, що ведуть до об'єднання і рекомбінації генетичного матеріалу у багатоклітинних і одноклітинних еукаріот, підіб'ємо деякі підсумки. Ці процеси дуже різноманітні зовні, але по суті своїй зводяться до закономірної зміни діплофази на гаплофазу в результаті мейозу і зміні гаплофази на діплофазу в результаті запліднення. Виняток становить парасек- суальних процес. Знання цих процесів необхідно при дослідженні кожного виду організму для правильного планування експерименту з генетичного аналізу і для інтерпретації його результатів. На цих знаннях грунтується велика галузь генетики - приватна генетика окремих видів.
Знання генетичного сенсу життєвих циклів має общебиологичне значення, оскільки дозволяє оцінити роль тих чи інших стадій розвитку організму в забезпеченні стабільності і комбинативной мінливості виду в природі.
Контрольні питання і завдання: