| Головна |
| «« | ЗМІСТ | »» |
|---|
У волоскових клітин немає аксона, вони з'єднані з відростками другого нейрона (біполярної клітини), тіло якого лежить в равлику. Слухові волокна володіють спонтанною активністю (за відсутності стимуляції). До волоскові клітини також йдуть еферентні волокна від клітин ядра оливи. Якщо їх стимулювати, то це призводить до пригнічення відповіді воло- Сковен клітин.
Слуховий тракт починається від первинних чутливих нейронів, які розташовуються в спіральному ганглії поблизу равлики. Аксони цих нейронів прямують до кохлеарним ядер середнього мозку, де здійснюється перша синаптичне перемикання. Аксонинейронів цих ядер направляються до ипси- і контралатеральной оліварним комплексам (Довгастий мозок), що забезпечує інтегрування сигналів від лівого і правого вуха на цьому рівні (менша частина волокон залишається в своєму півкулі, а велика частина переходить на протилежну сторону). Далі, після синаптичного перемикання в ядрі латеральної петлі, слуховий тракт проходить через нижні горбки четверохолмия і медіальне колінчаті тіло в первинну слухову кору (поле 41 по Бродману). Таким чином, слуховий тракт складається, принаймні, з п'яти нейронів (рис. 14.5).
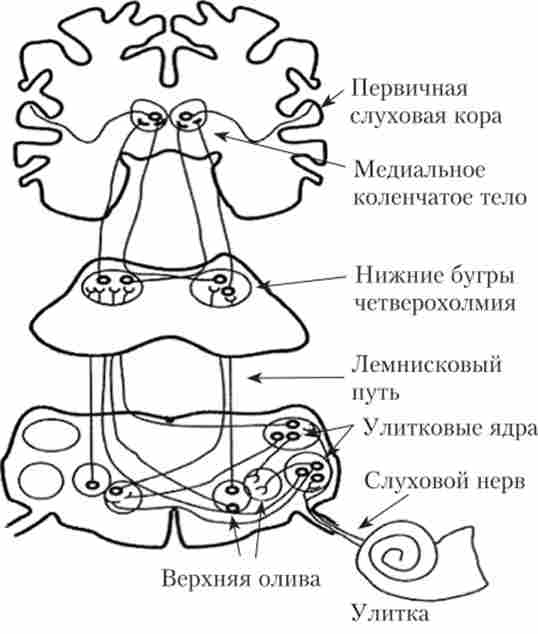
Мал. 14.5. Провідні шляхи і переключательние ядра слуховий системи1
Слуховий нерв містить порівняно мало волокон, але слух при цьому залишається гострий. Це пов'язано з двома факторами:
1) кожне слухове волокно, досягнувши кохлеарного ядра в стовбурі мозку, ділиться на безліч кінцевих гілок, ці гілки строго Тонотопична 1
розподіляються в кохлеарном ядрі (одна клітина має в кохлеарном ядрі множинне представництво);
2) в різних частинах кохлеарного ядра є різні типи синапсів і клітин. Головна частина містить вельми великі і характерні релейні нейрони, які служать морфологічним субстратом різних видів переробки слухової інформації.
На рівні підкіркових центрів слуху відбуваються процеси, що забезпечують можливість просторового (бинаурального) слуху. Для цього необхідне об'єднання інформації, одержуваної від правої і лівої равликів. Перший рівень об'єднання інформації від обох равликів - комплекс верхньої оливи. Він складається з латерального (5-образного) і медіального (акцессорного) ядер. Функція нейронів латерального ядра складається, в основному, в кодуванні частотних характеристик звукового сигналу. Нейрони відповідають в основному на іпсілатсральную стимуляцію. Медіальне (акцессорное) ядро отримує сигнали як від ипси-, так і від контрлатеральной равлики. Там є нейрони-детектори бінауральної тимчасової затримки, яка відображає напрямок локалізації джерела звуку.
Заднє двухолмие середнього мозку отримує сигнали від спинного кохлеарного ядра і латерального (акцессорного) ядра верхньої оливи. Отже, механізми просторового слуху в двухолміе дублюють механізми верхньої оливи і збагачуються новими детектуючих властивостями нейронів. Вперше тут з'являються нейрони, які відповідають тільки при переміщенні стимулу (детектори руху слухового сигналу).
Медіальне коленчатое тіло таламуса (МКТ) теж містить багато високо селективних детекторів локалізації та руху звуку. Крім того, звук у ряду нейронів МКТ крім реакції на включення звуку викликає ще один компонент - реверберірующій розряд, який виникає через секунди після вимкнення звуку і триває до декількох секунд. Це пов'язано з з'являтимуться тільки на цьому рівні механізмом фіксації на досить тривалий час інформації про локалізацію звуку в просторі.