| Головна |
| «« | ЗМІСТ | »» |
|---|
У процесі індивідуального розвитку організм закономірно змінює свої характеристики. Особливо інтенсивні зміни відбуваються в ембріональному періоді онтогенезу, коли з зиготи формуються структури багатоклітинного організму. При цьому все різноманіття клітин, що виконують в організмі різні функції, відбувається з однієї клітини шляхом мітотичного поділу. Так як в результаті мітозу дочірні клітини отримують повноцінну спадкову інформацію, укладену в каріотипі, всі клітини організму в генотипичному щодо рівноцінні. Деякі відмінності, однак, спостерігаються за рахунок цитоплазматичних генів, наприклад мітохондріальних, які розподіляються при розподілі не строго рівномірно.
Чим визначаються морфологічні, фізіологічні та біохімічні відмінності, що з'являються між клітинами в ході розвитку? В процесі овогенеза в цитоплазмі яйцеклітини накопичуються не тільки багаті енергією речовини, що забезпечують розвиток зародка, але і мРНК для синтезу білків, необхідних на самих ранніх стадіях ембріонального розвитку. Розподіл цих речовин в цитоплазмі яйцеклітини виявляється нерівномірним. Проникнення сперматозоїда в яйцеклітину викликає перерозподіл окремих компонентів в обсязі клітини, в зв'язку з чим вже при перших діленнях зиготи в дочірніх клітинах виявляється цитоплазма з різним складом речовин.
Взаємодія між компонентами цитоплазми і ядром призводить до дерепрессии визначених генів. Їх продукти визначають подальше поглиблення розходжень між різними частинами зародка, т. Е. диференціювання. Виникаючі розбіжності породжують нові взаємодії між сусідніми клітинними групами, які викликають дерепресія нових генів, внаслідок чого змінюється спектр активних генів і, отже, генетична програма на наступний відрізок процесу розвитку. Таким чином, в ході індивідуального розвитку спочатку репресований геном зиготи піддається поступової дерепрессии, причому в різних частинах зародка дерепрессіруются різні групи генів. Набір активно функціонують генів визначає своєрідність спектра білків, які синтезуються клітинами, які виконують різні функції.
Гени, транскрибуватися в ембріональному періоді, до моменту народження або безпосередньо після нього репресуються, в той же час активуються гени, що визначають специфічні функції клітин в дорослому організмі. Нерідко речовини, які продукують певним типом клітин в різні періоди онтогенезу, не однакові за своїми властивостями. Зміна властивостей диктується зміною умов існування організму, наприклад в ембріональному і постембріональному періодах розвитку. Ці відмінності пояснюються зміною функціонування близьких, але не ідентичних по укладеній в них інформації генів. Такі гени в ряді випадків утворюють групи, які отримали назву мультігенних сімейств. Прикладом цього є гени гемоглобіну.
Мул'тігенное сімейство - це група генів, дуже близьких по нуклеотидних послідовностей, з подібними фенотипичними функціями. Число генів в різних родинах у представників різних видів варіюється від одиниць до декількох сотень. Наприклад, число генів гістонів у різних видів в окремих родинах коливається від 10 до 1200, генів тРНК - від 6 до 400, генів 5SPHK- від 200 до 24 000, генів а-глобинов - від 1 до 5, (5-глобинов - від 2 до 7. до числа білків, кодованих мультігенних родинами, крім зазначених вище, відносяться актину і тубуліну, які відіграють важливу роль в рухливості клітин, колаген сполучної тканини, деякі білки клітинних мембран і сироватки крові.
Мультігенних сімейства можуть бути організовані в геномі по-різному (рис. 7.4). Так, члени сімейства ідентичних генів РНК у людини розташовуються у вигляді тандемних повторів, в яких структурні гени розділені некодуючими - спейсернимі - ділянками.
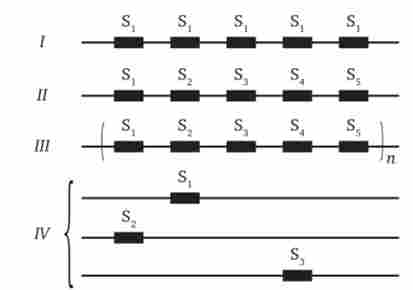
Мал. 7.4. Типи організації мультігенних сімейств.
I - ідентичні гени, зчеплені тандемно (гени рРНК); П - близькі, але не ідентичні гени, зчеплені тандемно (гени глобинов); III - скупчення неідентичних генів, зчеплених тандемно (гістонові гени); IV - близькі гени, розсіяні по декільком хромосомами (гени актину або тубулінів)
Однак гени сімейства не завжди ідентичні. Наприклад, в родинах глобінових генів тан- Демня зчеплені близькі, але не однакові гени.
У разі гістонових генів у деяких видів тандемно повторюються цілі пакети (кластери) неідентичних генів, що визначають синтез різних видів гістонів, що може бути описано формулою Н2А - НЗ - Н2В - Н4 - Н1 -. Можливо також дифузне розподіл генів сімейства за кількома хромосомами, що показано для генів актину і тубулінів.
Описані варіанти організації мультігенних сімейств створюють необхідні умови для ефективного регулювання експресії відповідних генів. Так, якщо продукт певного гена необхідний лише на невеликому відрізку часу в онтогенезі, але в значних кількостях, мультігенное сімейство утворено великим числом ідентичних генних копій, зазвичай з'єднаних тандемно. Прикладом можуть служити гени рРНК, які в геномі соматичних клітин дорослої шпорцевой жаби представлено 450 копіями. Разом з тим в овогенезі для швидкого освіти необхідної кількості рибосом, яких в яйці Xenopus міститься близько 1012, гени рРНК амплифицируют і число їх копій зростає в 4000 разів. В інших мультігенних родинах, що складаються, як правило, з неідентичних генів, в ході онтогенезу відбувається перемикання з одного гена на інший. Білки, контрольовані певними генами такого сімейства, найкращим чином відповідають або умовам на різних стадіях онтогенезу, або клітинам різних типів.
Найбільш вивчені в цьому відношенні мультігенних сімейства а- і Р-глобінових генів (рис. 7.5). У людини вони представлені кластером з 7 Р-глобінових генів, розташованих в 11-й хромосомі, і кластером з 5 а-глобінових генів, що локалізуються в 16-й хромосомі. У ембріонів людини активно функціонують t, (Дзета) -глобіновий ген з а-сімейства і е (епсилон) -глобіновий ген з Р-сімейства, що забезпечують освіту ембріонального |2е2-гемоглобіну.
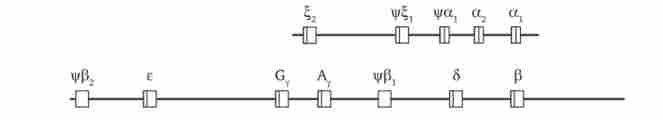
Мал. 7.5. Розподіл генів а- і Р-глобинов людини.
Глобінових гени зображені у вигляді прямокутників; вертикальні смуги всередині відповідають екзонних, інші ділянки - інтрони
На більш пізніх стадіях онтогенезу у плода ці гени репресуються, але дерепрессіруются інші гени сімейств, що визначають синтез фетального а2у2-гемоглобіну. Після народження починають експресувати 5 (дельта) - і р (бета) -глобіновие гени, що забезпечують освіту переважаючого а2Р2- і мінорного а282-видів гемоглобіну дорослої людини. В обох кластерах є також неекспрессірующіеся псевдогени раа, v / ^, vpPj, РР2.
Перемикання генів в мультігенних родинах відбувається не тільки у відповідності зі стадією індивідуального розвитку, але і з типом і місцем локалізації клітин в організмі. Так, ембріональний гемоглобін ^2е2 утворюється мають ядра Мегалобластов в стінці жовткового мішка. На 6-му тижні розвитку відбувається зміна експрессіруемих генів і перехід до синтезу гемоглобіну без'ядерними еритроцитами печінки і селезінки плода. Пізніше головним місцем утворення гемоглобіну стає кістковий мозок, де незабаром після народження починається синтез дорослих гемоглобинов а2(32 і а2б2. Зміна типів синтезованого гемоглобіну у ембріона, плода і після народження пов'язана з конкретними умовами існування організму на різних стадіях онтогенезу. Так, у людини гемоглобін плода має більш високу спорідненість до кисню, ніж гемоглобін дорослого, що полегшує перенесення кисню через плаценту.
Таким чином, зміна характеристик фенотипу організму на різних стадіях онтогенезу є результатом регуляції експресії генів, яка має на меті в одних випадках нарощування продукції певних білків, а в інших - перехід від синтезу одного білка до синтезу білка більше відповідного постійно змінюваних умов існування.