| Головна |
| «« | ЗМІСТ | »» |
|---|
Таламус є масивне парне освіту (рис. 11.1), що містить близько 120 ядер сірої речовини.
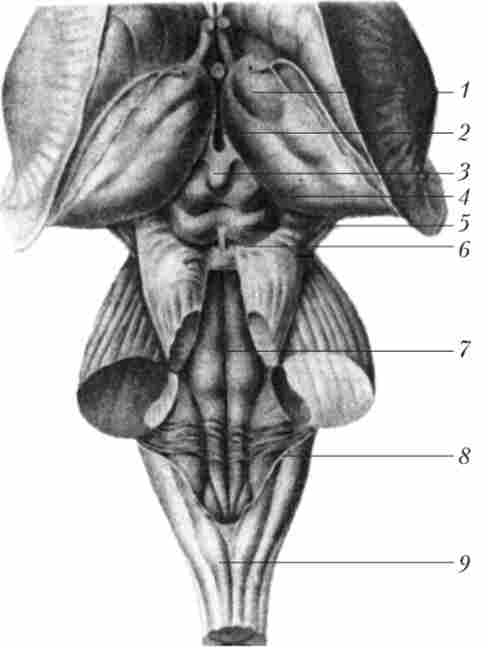
Мал. 11.1. Стовбур мозку і таламус1:
Діяльність таламуса тісно пов'язана з аналізом аферентних сигналів, з регуляцією функціонального стану організму. Він взаємодіє з корою б. п.
Таламус включає в себе власне зоровий бугор, потім метаталамус (медіальні і латеральні колінчаті тіла) і подушку.
За морфологічними критеріями всі ядра таламуса об'єднують в шість груп:
За функціональної ролі в діяльності нервової системи в таламусі виділяють ядра специфічні, неспецифічні і асоціативні (рис. 11.2).
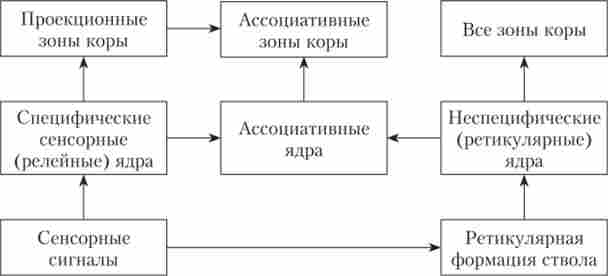
Мал. 11.2. Схематичне зображення основних груп ядер таламуса і їх зв'язків
Специфічні ядра таламуса. Для специфічних ядер таламуса характерні наступні особливості. Ці ядра мають локальну проекцію в строго певні зони кори. Вони моносінантічно пов'язані з нейронами 3-го і 4-го шару кори. У специфічних ядрах перемикається аферентна пульсація від сенсорних ядер нижчих структур. Основну масу клітин складають так звані релейні (Переключательние) клітини. Роздратування будь-якої рецепторной області спочатку викликає у відповідному специфічному ядрі відповідну реакцію у вигляді потенціалу, а потім роздратування самого ядра викликає відповідну реакцію вже в певній корковою зоні. У кожному ядрі виражена топична організація, т. Е. Кожної ділянки шкіри, сітківки і т. Д. Відповідає певна зона таламуса.
Слухова система проектується в медіальні колінчаті тіла (МКТ), Які є предкорковим рівнем аналізу слухових сигналів. На одні й ті ж нейрони медіального колінчастого тіла можуть конвергировать збудження від багатьох нейронів заднього двухолмия середнього мозку.
Зорова сенсорна система на рівні таламуса представлена латеральний колінчатими тілами (ЛКТ). Вони вважаються найбільш складно організованими специфічними ядрами таламуса. Від них волокна йдуть в 17-е і 18-е поля кори (потилична область).
Крім сенсорних, до релейних ядер таламуса відносяться ще моторні ядра і ядра передній групи. Це єдиний комплекс. В моторних ядрах перемикається афферентация, що йде від ядер мозочка, блідої кулі, вестибулярних і пропріореценторов до моторної кори.
Релейная функція ядер передньої групи полягає в перемиканні імпульсів від маміллярних тел гіпоталамуса до лімбічної системи. Ядра передньої групи відносять до лімбічної системи (коло Пайпеца).
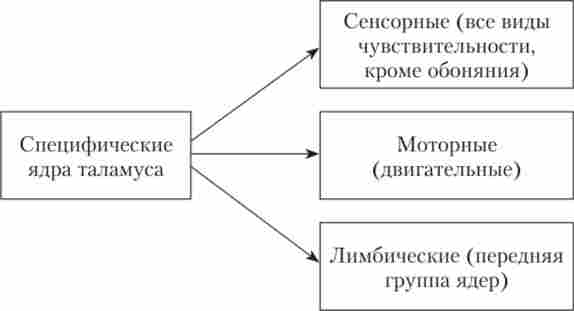
Мал. 113. Групи специфічних ядер таламуса
Таким чином, специфічні ядра являють собою найважливішу частину основних сенсорних і моторних систем, а руйнування релейних ядер призводить до повної і незворотної втрати відповідної чутливості або порушень рухів.
Неспецифічні ядра таламуса. Вони не належать до певної сенсорної або моторної системі, морфологічно і функціонально вони пов'язані з багатьма системами і беруть участь разом з РФ в здійсненні неспецифічних функцій. Нейронні мережі цих ядер мають ретикулярное будова: густа мережа нейронів з довгими слабо розгалуженим ден- ДРІТ.
Зв'язок у неспецифічних ядер з корою в основному полісинаптичних, волокна йдуть до всіх верств кори. Проектуються вони в кору більш дифузно, ніж специфічні. Аферентна інформація надходить до них в основному з РФ, а також з гіпоталамуса, лімбічної системи, базальних гангліїв, специфічних ядер таламуса. Неспецифічні ядра отримують сигнали і від специфічних ядер.
Як правило, одиночне електричне роздратування цих ядер не викликає одиночній відповідної реакції кори. Ритмічна низькочастотна стимуляція призводить до реакції синхронізації біоелектричної активності головного мозку, а високочастотна - до десинхронізації (реакція активації кори). Ця реакція реєструється в неспецифічних зонах кори, так як в специфічних вона пригнічується специфічної импульсацией.
Неспецифічні ядра таламуса надають на кору модулюючий вплив, т. Е. Регулюють її функціональний стан. Вони змінюють її реактивність до специфічних сигналів. Так само як і у випадку з РФ, діяльність неспецифічної таламичної системи тісно пов'язана з механізмами розвитку сну, саморегуляції функціонального стану і вищої нервової діяльності (ВНД).
Специфічна і неспецифічна системи таламуса знаходяться у взаємодії один з одним. Так, було показано, що якщо неснеціфічна система підсилює специфічну, то специфічна пригнічує несіеці- фичного.
Асоціативні ядра таламуса. Це найбільш пізно диференціюються в еволюції, але і найбільш активно розвиваються відділи таламуса. Волокна від цих ядер направляються в основному до асоціативних областей кори і, частково, до специфічних проекційним областям. Зв'язки з корою в основному моносинаптічеськие. Основні аферентні сигнали надходять від інших ядер таламуса, а нс з периферії.
Електричне подразнення асоціативних ядер таламуса викликає відповідні реакції в асоціативних областях кори. Багато з цих ядер здатні реагувати на подразнення різних сенсорних входів, деякі взагалі реагують тільки на комплексні подразнення. У них можуть взаємодіяти збудження різних сенсорних систем, т. Е. Вони здійснюють інтеграцію імпульсації від всіх сенсорних систем.
Крім передачі проекційних впливів на кору, нейрони таламуса самі можуть здійснювати замикання рефлекторних шляхів без участі кори і таким чином здійснювати самостійно складні рефлекторні функції.
Таламус часто називають вищим центром больової чутливості. Він здійснює аналіз больових сигналів і організовує больові відповідні реакції. Імпульси, що йдуть до нейронам таламуса від пошкоджених ділянок тіла, активують ці нейрони і викликають больові відчуття. Таким чином, больові відчуття пов'язані з порушенням неспецифічних нейронів таламуса, для цього не обов'язково участь кори. У корі формується вже суб'єктивне ставлення до больового стимулу.