| Головна |
| «« | ЗМІСТ | »» |
|---|
Існує кілька підходів до пояснення причин виникнення сну:
Першою на початку XX ст. сформувалася судинна (циркуляторная) теорія сну. Її прихильники пов'язували сої зі зміною кровотоку в мозку. Одні пов'язували сон з відтоком крові та анемією (К. Бернар, А. Моссо, І. Р. Тарханов), інші - з припливом крові до головного мозку і гіперемією (Броун, Шлейх і ін.). За сучасними даними (отриманим методом реоенцефалографії) виявилося, що під час сну відбуваються коливання кровонаповнення судин мозку.
перші гуморальні (хімічні) теорії сну пояснювали наступ сну за рахунок речовин, що накопичуються в організмі під час діяльності: збіднення клітин киснем, накопичення вуглекислого газу, молочної кислоти, холестерину і т. д. Р. Лежандр і X. Пьерон (1910) вважали, що сон настає в результаті накопичення токсичних речовин (продуктів обміну). Вони експериментально показали, що якщо одному собаці довго не давати спати, а йотом її кров або ліквор перелити інший собаці, то у другій наступав сон. Також вони вивчали мозкову тканину довго не спали тварин і виявили серйозні зміни: змінилася форма ядер, мембрана була поїдена. Складалося враження, що тканина була вражена якоюсь отрутою. В результаті виникло поняття «сонний отрута», або гипнотоксин, на роль якого було багато кандидатів (див. Вище). Однак гуморальним теоріям суперечили спостереження за зрослими близнюками. П. К. Анохін близько року спостерігав за зрослими грудною кліткою сіамськими близнюками (отже, у них була спільна система кровообігу). Досить часто він бачив картину, при якій одна голова спала, а інша - ні. Проте в 1965 р швейцарському нейрофізіологу А. Моньє на тваринної моделі сіамських близнюків вдалося частково підтвердити теорію про гипнотоксинов і навіть виділити з крові тварин особливий поліпептид, який виділявся в кров довго не спали тварин. Якщо його ввести іншій тварині, то воно миттєво засипало. Але різний час сну у сіамських близнюків, за якими спостерігав П. К. Анохін, і у інших залишається фактом. Таким чином, гипнотоксин може бути одним з механізмів, але не єдиним механізмом сну.
В результаті вивчення різних відділів мозку, їх стимуляції або руйнування з'явилися нервові теорії сну. У 1924 р швейцарський фізіолог В. Р. Гесс проводив досліди з введенням електродів в головний мозок. Дратуючи гіпоталамус, він викликав у тварин сон. Після його експериментів і ряду спостережень утвердилася думка про існування особливих центрів в головному мозку - так званих центрів сну. Однак І. П. Павлов вважав, що сон виникає з іншої причини. Він спостерігав за собаками при виробленні умовних рефлексів і виявив, що сон у них виникав в результаті гальмування кори б. п. Він і його послідовники вважали, що сон являє собою пасивний процес, який виникає в результаті припинення потоку інформації до кори. Отже, на сьогоднішній день нервові теорії сну прийнято ділити на активні і пасивні теорії (табл. 7.1). активні теорії виходять з того, що сон виникає в результаті активного процесу - збудження певних ділянок мозку (центрів сну), а це, в свою чергу, призводить до засипання. прихильники пасивних теорій вважають, що сон виникає в результаті припинення надходження сигналів (інформації) до мозку, т. е. пасивно. Їх ще називають теоріями деафферентации. Тобто «заснути» і «не зуміти залишитися бодрствующим» - це різні речі.
Існують експериментальні підтвердження як активних, так і пасивних теорій.
Таблиця 7.1
Порівняння пасивних і активних теорій сну
|
активні теорії |
пасивні теорії |
|
Гесс показав, що слабке електричне роздратування чітко обмеженою області проміжного мозку кішки викликало у неї сон з усіма підготовчими стадіями |
У клінічних дослідженнях було показано, що сон може викликати пухлину середнього мозку, яка здавлює аферентні шляхи і припиняє потік інформації в кору б. п. |
|
Відкрито інші структури, роздратування яких може викликати сон: хвостате ядро, неснеціфічні ядра таламуса, преоптична область гіпоталамуса |
І. 11. Павлов розглядав сон як результат гальмування кори б. п. Він вважав, що якщо виключити потік імпульсів в кору, то настане гальмування і сон |
|
" |
Експерименти Бремера з перерізанням на різних рівнях висхідних шляхів до кори (деафферентаціі) |
|
- |
Відкриття Мегун і Моруцци висхідній активує системи мозку (РФ) |
У 1935 р бельгійський нейрофізіолог Бремер провів досліди з перерізанням аферентних шляхів головного мозку на різних рівнях. У першому випадку він перерізав висхідні шляху на рівні першого шийного хребця, як би від'єднавши головний мозок від спинного (препарат отримав назву ізольований мозок - Encephale isole) (Рис. 7.1). В цьому випадку у кішки була присутня нормальна зміна періодів сну і неспання.
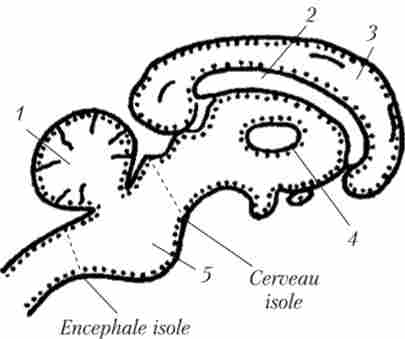
Мал. 7.1. Ілюстрація експериментів Бремера з перерізання висхідних шляхів у мозку кішок:
1 - мозочок; 2 - мозолисте тіло; 3 - кора; 4 - таламус; 5 - варолиев міст
Перерізавши ж шляху в кору б. п. на рівні середнього мозку, він отримав препарат, який назвав Cerveau isole - ізольовані півкулі. В цьому випадку спостерігалася і поведінкова, і електрофізіологічних картина сну. Сон в останньому випадку пояснювали зниженням припливу інформації до кори (так як шляху були перерізані). Однак виникло питання, звідки йшли ці сигнали. Відповідь була отримана в результаті робіт американця Мегун і італійця Моруцци в 1949 р, коли вони показали, що між місцями перерезок в експериментах Бремера розташована ретикулярна формація. У ній знаходяться нейрони, які формують висхідні активують впливу на кору і дозволяють їй підтримувати певний рівень неспання, перебувати в тонусі.
Пізніше з'ясувалося, що в мозку працюють кілька центрів сну і центрів неспання. До центрам сну відносять ядра шва стовбура, деякі ядра ретикулярної формації, преоптічної область гіпоталамуса. Це ті центри, роздратування яких викликає засипання (рис. 7.2).
Роздратування блакитного плями стовбура, чорної субстанції, деяких ядер моста і заднього гіпоталамуса викликає активацію і пробудження, т. Е. Підвищення рівня неспання.
П. К. Анохін запропонував свій варіант єдиної теорії сну, що об'єднує активні і пасивні теорії. Він вважав, що дійсно існують гипоталамичні центри сну, але під час неспання вони знаходяться під постійним тонічним гальмівним впливом з боку кори б. п. Якщо робочий тонус коркових нейронів знижується (в результаті зменшення потоку інформації в кору або її стомлення), то їх гальмівний вплив на нижні центри послаблюється. Гіпоталамічні центри як би вивільняються з-під гальмівного впливу кори і викликають вегетативні та інші зміни, характерні для стану сну. Вони також пригнічують висхідні активуючі впливи з ретикулярної формації на кору. Ці зміни представляються циклічними. В якості ілюстрації цієї теорії Анохін запропонував розглянути сон новонародженого і його зміни в онтогенезі. Так, у новонародженого сон становить від 16 до 20 год на добу, м. Тобто він досить тривалий. Це пов'язано з тим, що кора у новонародженого ще дуже незріла і не може довго підтримувати неспання. Зате гипоталамичні і стовбурові центру сну досить дозріли. У міру дозрівання кори тривалість сну швидко зменшується, кора все довше і довше може не спати. В результаті сон стає монофазним - один раз на добу і за тривалістю становить близько 8 год (параграф 7.4.).
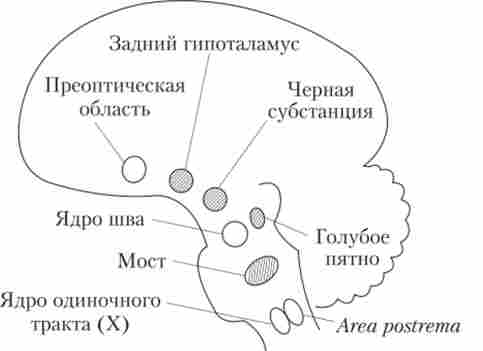
Мал. 7.2. Звернення до центру сну і неспання в головному мозку1
Існує також і інформаційна теорія Н. Вінера, яка швидше за пояснює не чому, а для чого виникає сон. Вінер пропонує наступне пояснення виникнення сну. Протягом дня мозок накопичує величезну кількість інформації, яка повинна бути або перенесена в довгострокову пам'ять, або стерта. Ночио під час сну частина інформації переноситься з короткочасної пам'яті в довготривалу. При цьому особливості переробки інформації вимагають відключення від зовнішнього світу.