| Головна |
| «« | ЗМІСТ | »» |
|---|
Здатність гомойотермних тварин підтримувати постійну температуру тіла незважаючи на різкі коливання температури навколишнього середовища - одне з найважливіших біологічних регуляторних явищ. Процеси терморегуляції протікають незалежно від свідомості і не припиняються під час сну. Початковим ланкою терморегулюючої системи шкіри є численні терморецептори. Оскільки зміни температури навколишнього середовища створюють суб'єктивні відчуття тепла або холоду, то можна припустити, що в шкірі є відповідно теплові та холодові рецептори. Дійсно, в перших же об'єктивних дослідженнях терморецепціі шкіри, проведених на початку 20-х років XX ст., Були виявлені особливі точки, в яких при нагріванні або охолодженні відбувалося збільшення частоти аферентних потенціалів дії в нервових волокнах, що іннервують ці ділянки. За реакцією на адекватний стимул температурні рецептори були схожі на медленноадаптірую- щіеся механорецептори шкіри (рис. 15.9). У спокої у обох типів терморецепторов була зареєстрована спонтанна імпульсна активність. У відповідь на ступенчатообразное зміна температури (підвищення або зниження) частота аферентних потенціалів дії швидко підвищувалася, досягаючи максимального значення, і потім порівняно швидко знижувалася до певного рівня, а далі процес уповільнювався. Дія температурних стимулів з різними швидкостями наростання дозволяє віднести
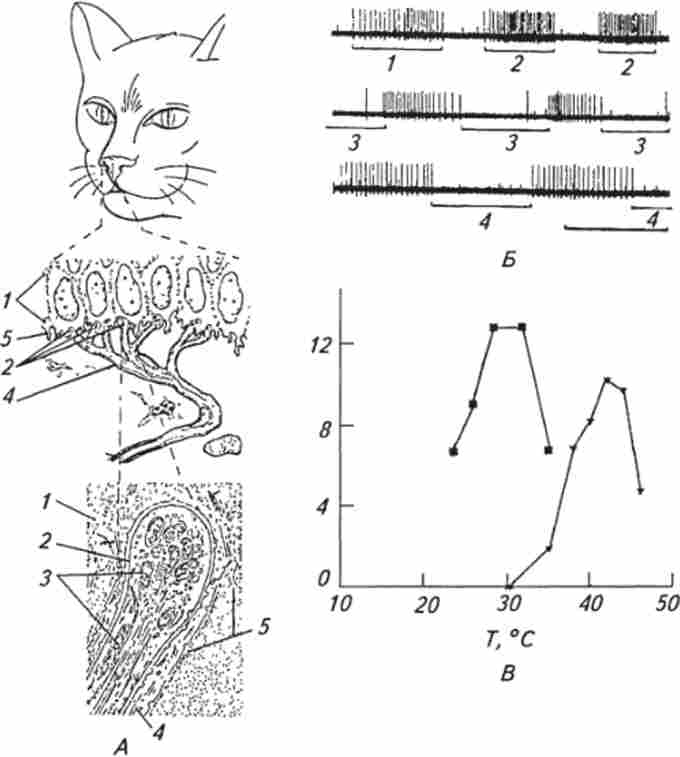
Мал. 15.9. Терморецептори шкіри:
A. Схематичне зображення одиночного холодового рецептора на безволосої шкірі носа кішки, крапка - місце його локалізації: / - епідерміс; 2 - рецепторное закінчення; J - мітохондрії всередині рецепторного закінчення; 4 - шванновскими клітина; 5 базальнамембрана. Б Імпульсна реакція холодового рецептора шкіри носа кішки на температурні стимули (внизу під електричною реакцією горизонтальними прямими відзначено дію температурного стимулу): / - обдування охолодженим повітрям. 2-дотик охолодженим термодом (мідна зволікання діаметром 60 мкм); 3 - дотик до ділянки шкіри з холодових рецептором нагрітим термодом; 4 обдування шкіри нагрітим повітрям. горизонтальна лінія внизу під електричними відповідями холодового рецептора - калібрування часу I с.
B. Залежність між середньою частотою імпульсної активності холодового (квадрати) і теплового (трикутники) рецепторів шкіри і температурою. Охолодження шкіри нижче 35 * С у холодового рецептора викликає підвищення імпульсної активності в афферентном волокні, а у теплового - припинення і, навпаки, при підвищенні температури вище 35 * С імпульсна активність холодового рецептора пригнічується, а теплового підвищується; по осі ординат - частота потенціалів дії, імп / с, а по осі абсцис - температура, С
температурні рецептори до детекторам амплітуди швидкості. Дослідження залежності частоти аферентних потенціалів дії від величини температури показало, що середня частота імпульсної активності при нагріванні або охолодженні зростає до максимального значення, а потім в обох випадках починає зменшуватися до повного зникнення. Таким чином, співвідношення носить куполоподібний характер (див. Рис. 15.9, В).
Які ж морфологічні кореляти теплових і холодових рецепторів? З кінця XIX в. по 60-ті роки XX ст. існувала думка, засноване на суб'єктивних даних досліджень на людині, що холодових рецепторів відповідають інкапсульовані тільця Краузе, що нагадують за своєю будовою тільця Пачіно, а тепловим - тільця Руффини. Однак об'єктивні дослідження з використанням реєстрації афферентной імпуль- сации показали, що ці рецептори є шкірними механорецепторами (див. Попередній розділ). Експерименти з одночасною реєстрацією імпульсної активності і морфологічної маркуванням виявили, що терморецепторами відповідають не- інкапсульовані нервові закінчення, локалізовані в обох шарах шкіри. Наприклад, холодовий рецептор безволосої частини носа кішки (див. Рис. 15.9, Л) є неразветвлен- ве неміелінізірованнимі закінчення в епідермісі діаметром
2,5 ... 4 мкм; розширений його кінчик заповнений дрібними мітохондріями. Терморецептори утворені тонкими міелінізіро- ванними нервовими волокнами діаметром 1,5 ... 5 мкм.
Зміни частоти аферентних потенціалів дії в нервових волокнах терморецепторов має передувати зміна мембранного потенціалу афферентной області (рецепторного потенціалу) (див. Рис. 15.1). Відомо, що формування мембранного потенціалу обумовлено пасивним і активним іонним транспортом. Величина пасивної компоненти мембранного потенціалу обчислюється за допомогою рівняння Голдмана. Тому можна припустити, що варіації в температурі будуть безпосередньо змінювати пасивний транспорт іонів і відповідно впливати на амплітуду мембранного потенціалу. Однак прості розрахунки показують, що коливання температури на 10 ° С змінює мембранний потенціал всього лише на 2 мВ. Якщо врахувати, що терморецептори мають спонтанної активністю, т. Е. Мембранний потенціал знаходиться в області порога генерації потенціалів дії, то даної величини зміни мембранного потенціалу все ж буде недостатньо для помітного збільшення частоти імпульсації. Потрібно відзначити, що у деяких видів тварин теплові рецептори мають надзвичайно високу чутливість. Так, в лицьовій ямці гримучої змії знаходяться розгалужені нервові закінчення, що реагують на підвищення температури всього на 0,002 ° С. Дослідження останніх років дозволяють вважати, що в рецепторной мембрані є канали, які активуються підвищенням або пониженням температури. Додатково, що стосується рецепції холоду, то великий експериментальний матеріал вказує на участь в цьому процесі активного іонного транспорту - іонного насоса.
Зокрема, передбачається, що в холодових рецепторах при звичайній температурі тіла тварини мембранний потенціал в рецепторних закінченнях зрушать убік гіперполяризації Електрогене іонним насосом. Робота іонної помпи залежить від температури, тому її зниження, охолодження нервової тер- минали буде загальмовує роботу помпи, що викличе деполяризацію мембрани і відповідно збільшення частоти аферентних потенціалів дії.
Сукупність теплових і холодових рецепторів забезпечує функціонування системи терморегуляції організму тварини. Так, інтенсивне охолодження тіла збуджує холодові рецептори. За допомогою автономної нервової системи вони викликають звуження шкірних кровоносних судин, що зменшує інтенсивність кровотоку в шкірі і зберігає теплоту всередині організму. Зниження тепловіддачі досягається також зменшенням поверхні тіла (згортання в клубок) і зміною положення волосся або пір'я на тілі тварини. Одночасно через систему рухових нервів збільшується виділення теплоти в м'язах за рахунок напруги м'язів і м'язової тремтіння, а також за допомогою «розігрівають» рухів. Теплопродукція зростає, крім того, завдяки посиленню обміну речовин у внутрішніх органах.
При зовнішньому перегріванні тіла тварини теплові рецептори шкіри викликають процеси, зворотні тим, що відбувається при переохолодженні: розширення кровоносних судин, посилення потовиділення, почастішання дихання.