| Головна |
| «« | ЗМІСТ | »» |
|---|
1. Клітинна будова серцевого м'яза відрізняється від будови скелетних м'язів, хоча серцевий м'яз теж відноситься до поперечно-смугастим. Скелетні м'язи складаються з волокон, які є анатомічними сііцітіямі, т. Е. Багатоядерними структурами, що утворилися в результаті злиття одноядерних клітин-попередниць. Серцевий м'яз теж складається з волокон, але вони є функціональними синцитій, т. Е. В них клітини зберігають самостійність, хоча і працюють спільно. М'язові клітини серця контактують один з одним за допомогою вставних дисків, утворених клітинними мембранами. У вставних дисках знаходяться щілинні контакти (Нексус), що представляють собою, по суті справи, електричні синапси (рис. 6.4). Завдяки Нексус імпульси, що запускають скорочення серця, безперешкодно поширюються від однієї м'язової клітини до іншої.
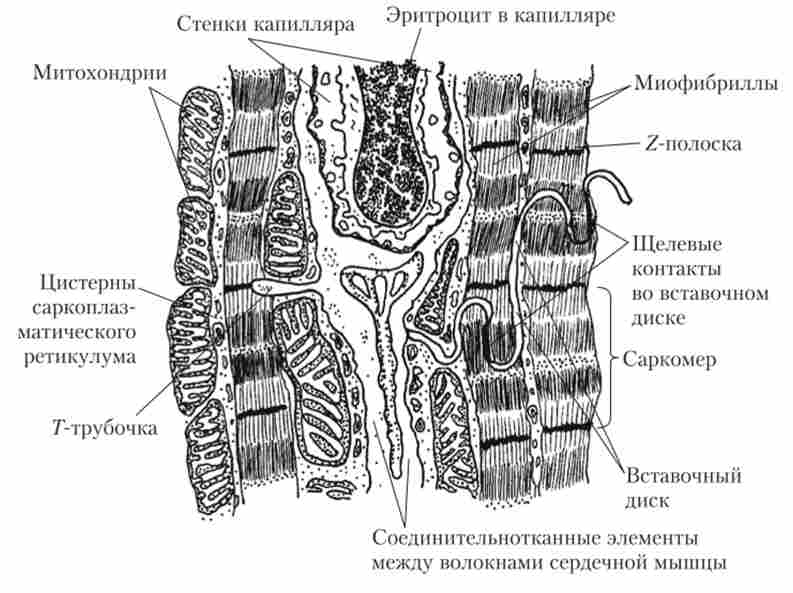
Мал. 6.4. Мікроскопічна будова серцевого м'яза (малюнок з мікрофотографії)
Багато клітини серцевого м'яза мають вирости, за допомогою яких вони вступають в контакт з клітинами сусідніх волокон (рис. 6.5). Завдяки цій особливості будови функціональним сінцітіем стає вся серцевий м'яз, а не одне волокно. Вважається, що міокард складається з двох функціональних сінцітіев: предсердного і шлуночкового.
Важливим анатомічним відмінністю серцевого м'яза від скелетної є більш висока каніллярізація, що вказує на дуже активне споживання кисню (див. Рис. 6.5). Це підтверджується присутністю в клітинах серцевого м'яза великої кількості мітохондрій. Їх щільність в серцевому м'язі в кілька разів вище, ніж навіть у швидкій скелетної м'язі (див. Рис. 6.4).
Автомати серця. Серед основної маси клітин серцевого м'яза, які утворюють робочий міокард, зустрічаються клітини, які мають менш
розвинений скорочувальний апарат. Їх основною функцією є нс скорочення, а генерація і проведення електричних сигналів. У стінці серця ці клітини утворюють єдину систему, яка отримала назву провідної системи серця.
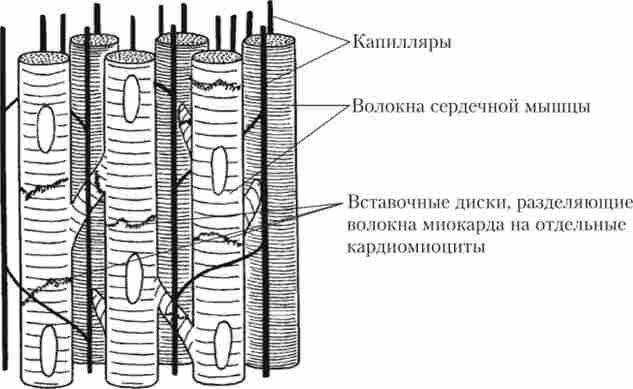
Мал. 6.5. Клітинна структура міокарда і розташування капілярів, що забезпечують його кровопостачання
Провідна система серця складається з клітин, що володіють здатністю мимовільно, без зовнішніх впливів, генерувати електричні імпульси з відносно постійною частотою. Ця здатність називається автоматией і пов'язана з присутністю в клітинних мембранах особливих типів іонних каналів (див. Нижче). Розташування в серце основних елементів провідної системи і частота генеруючих ними імпульсів показані на рис. 6.6.
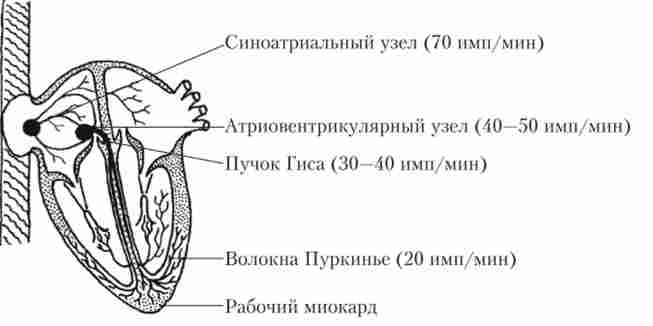
Мал. 6.6. Основні елементи провідної системи серця і частота, з якою вони здатні самостійно генерувати імпульси
З найбільшою частотою електричні імпульси генерує скупчення клітин провідної системи, розташоване в стінці правого передсердя в області гирла порожнистих вен. Ця структура називається синоатріальна вузлом автомашин. Генеруючи імпульси приблизно 70 раз / хв, синоатріальний вузол пригнічує автоматік) інших структур провідної системи і в нормі визначає ритм серцевих скорочень. В силу цього синоатріальний вузол називають головним водієм серцевого ритму (Пейсмейкер 1-го порядку). Від нього відходять пучки волокон провідної системи, що передають збудження до лівого передсердя (пучок Бахмана), правого передсердя і потім - до іншого вузла автоматии (пучки Венкебаха і Тореля). Цей вузол розташовується в нижній частині правого передсердя поблизу міжпередсердної перегородки і називається атріовентрикулярним. При блокаді синоатріального вузла він здатний генерувати імпульси з частотою 40-50 раз / хв (пейсмейкер 2-го порядку).
В атріовентрикулярному вузлі бере початок велике скупчення волокон провідної системи - пучок Гіса, за яким збудження передається від передсердь до шлуночків. Передсердя і шлуночки розділені невозбудимой сполучнотканинною перегородкою, і пучок Гіса є єдиною зв'язує їх проводить структурою. Увійшовши в шлуночки, пучок Гіса ділиться на праву і ліву ніжки, які йдуть по міжшлуночкової перегородки і потім розгалужуються на волокна Пуркіньє, передають порушення на робочий міокард.
І пучок Гіса, і ніжки пучка, і волокна Пуркіньє володіють не виявляється в нормі автоматичної активністю, що дозволяє віднести їх до пейсмейкер 3-го порядку. Прояв активності цих пейсмейкерів часто є причиною серцевих аритмій.
Природа автоматии серця пов'язана з присутністю в мембранах клітин провідної системи особливих білків: повільних натрієвих каналів (/ -каналів) і повільних кальцієвих каналів. Завдяки / -каналами через мембрану йде безперервний вхідний струм іонів Na+, який викликає повільну деполяризацію. Їй також сприяє поступове збільшення проникності мембрани для іонів Са2+. Таким чином, у пейсмей- Керн клітин відсутній «класичний» ПП, його замінює повільна деполяризація. Вона починає розвиватися з рівня -55 мВ (нижче значення мембранного потенціалу пейсмейкерних клітин не опускається) і завершується після досягнення критичного рівня деполяризації (-40 мВ) розвитком ПД.
ПД пейсмейкерной клітини формується за рахунок активації Са2+-кана- лов. Швидкі №+-канали в його формуванні не беруть участь (на відміну від нейронів, волокон скелетного м'яза і робочого міокарда), так як при мембранному потенціалі від -40 до -55 мВ вони інактивовані. На піку ПД (близько 0 мВ) відбувається інактивація Са2+-каналів і активація Ю - каналів. Результатом цього стає реполяризация, т. Е. Повернення мембранного потенціалу до рівня, з якого починалася повільна натрієва деполяризация (-55 мВ). Це призводить до активації / -каналів, і описана послідовність подій повторюється. Частота генерації потенціалів пейсмейкерной кліткою, таким чином, буде залежати
від швидкості розвитку повільної натрієвої деполяризації, яка визначається активністю повільних натрієвих каналів. ПД пейсмейкер- ної клітини серця і визначають його фази іонні струми зображені на рис. 6.7.
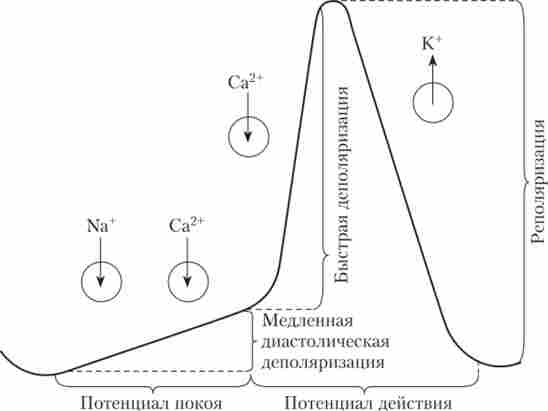
Мал. 6.7. Іонні потоки, що визначають різні фази потенціалу дії пейсмейкерной клітини:
/ У) - потік іонів в клітину; (Т) - потік іонів з клітки
Повільні натрієві канали є мішенню для регуляторних впливів ВНС. Симпатичні волокна виділяють НА, який через (3-адренорецептори і систему білків-посередників збільшує концентрацію цАМФ в клітці. Це призводить до зростання числа відкритих / -каналів, збільшення швидкості діастолічної деполяризації і частоти серцевих скорочень. Парасимпатичні волокна через медіатор АХ і М-холінорецептори надають на рівень цАМФ і ритм серця протилежне, т. е. гальмівне, дія (рис. 6.8).
Поширення збудження по провідній системі серця починається з синоатріального вузла, який пов'язаний з атріовентрикулярним вузлом міжвузловими пучками, а з лівим передсердям - міжпередсердної пучком. Швидкість проведення імпульсу в цих пучках становить близько 1 м / с (для порівняння: швидкість проведення імпульсу в волокнах робочого міокарда не перевищує 0,3 м / с). Порушення досягає атріовентрикулярного вузла за 0,03 с, після чого його поширення гальмується. Ця затримка дозволяє предсердиям перекачати кров в шлуночки до початку їх скорочення і називається атріовентрикулярної. Вона становить в сумі 0,13 с (0,09 с доводиться на вузол, а 0,04 с - на пучок Гіса). Далі збудження поширюється по ніжках пучка Гіса і волокнам Пуркіньє зі швидкістю від 1,4 до 4 м / с. Така велика швидкість досягається за рахунок
збільшення кількості щілинних контактів і їх проникності для іонів у вставних дисках волокон Пуркіньс. Висока швидкість поширення дозволяє порушення одночасно охопити всю масу волокон міокарда шлуночків і синхронізувати їх скорочення. Динаміка поширення збудження по серцю показана на рис. 6.9.
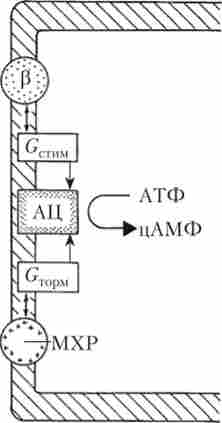
Мал. 6.8. Мембранний механізм, що забезпечує взаємодію симпатичних і парасимпатичних впливів на діяльність серця:
Р - p-адренорецептор; GCTllM - G-білок, що стимулює активність АЦ; АЦ - аденилатциклаза, яка виробляє цАМФ з АТФ; Згорм - G-білок, гальмуючий активність АЦ; МХР - М-холінорецептор
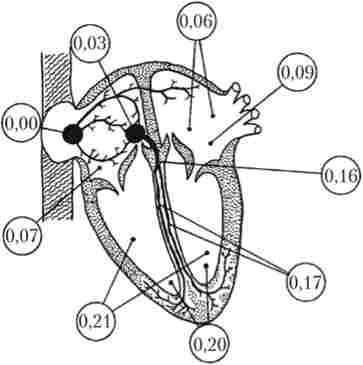
Мал. 6.9. Проведення збудження по серцю:
дано час появи ПД в різних ділянках міокарда в частках секунди від початку його створення в синоатріальної вузлі автоматии
Таким чином, провідна система серця забезпечує не тільки генерацію 11Д в стійкому ритмі, а й поширення збудження по серцевому м'язі зі швидкістю, що дозволяє серцю найкращим чином виконати свою насосну функцію. Однак безпосередньо в процес скорочення міокарда клітини провідної системи вносять дуже незначний внесок. Провідна роль в скороченні належить клітинам робочого міокарда.
ПД клітин робочого міокарда по амплітуді, тривалості, динаміці і іонної природі відрізняється як від ПД волокна скелетного м'яза, так і від ПД пейсмейкерних клітин (рис. 6.10).
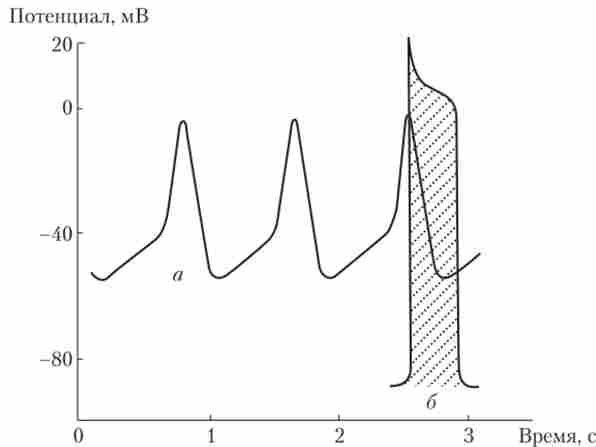
Мал. 6.10. Потенціали дії пейсмейкерних клітин (А) і клітини робочого міокарда (Б):
спостерігаються відмінності за амплітудою, тривалості і формі ПД
Для клітин робочого міокарда характерний ПП, що становить -85 мВ; основний внесок в його підтримку вносять іони К+. ПД починається з дуже швидкою деполяризації, викликаної активацією двох типів іонних каналів: швидких натрієвих (як в скелетному м'язі) і повільних кальцієвих (як в пейсмейкерних клітинах). Основний внесок в деполяризацию мембрани вносять іони Na+, що входять в клітину. Са2+-канали відрізняються тим, що вони повільно відкриваються, і тому іони Са2+ «Не встигають» взяти участь у розвитку деполяризації. При цьому Са2 - канали довго залишаються відкритими (0,2-0,3 с). На піку ПД (+20 мВ) швидкі Ка+-канали інактивуються, але потік іонів Са2 f через ще відкриті повільні Са2+-канали гальмує реполяризацию, формуючи плато (такого немає ні в скелетного м'яза, ні у пейсмейкерних клітин). Інактивація повільних Са2 ^ -Каналів і зростання проникності мембрани для іонів К+ повертають мембранний потенціал до рівня спокою (фаза реполяризації). Форма ПД клітини робочого міокарда і іонні потоки, що визначають його фази, показані на рис. 6.11.
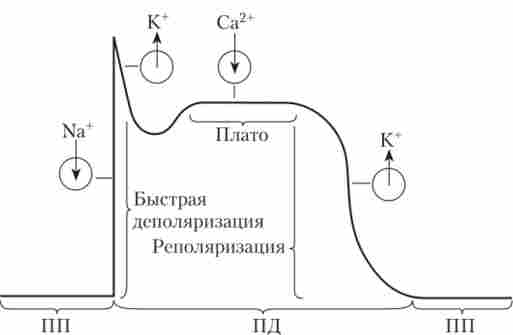
Мал. 6.11. Іонні потоки, що визначають різні фази потенціалу дії клітини робочого міокарда:
^ - потік іонів в клітину; ф - потік іонів з клітки
Як і всі збудливі клітини, клітини робочого міокарда мають реф- характерних, т. Е. Несприйнятністю до повторних стимулам під час розвитку ПД. Період повної несприйнятливості називається абсолютної рефрактерност'ю, а період, коли під дією сильного подразника можна отримати зменшений відповідь, - відносної рефрактерност'ю (Рис. 6.12). У нормі тривалість ПД клітин робочого міокарда шлуночків становить 0,3 с (передсердь - ОД с), період абсолютної рефрактерності дорівнює приблизно 0,27 с, а відносної - 0,03 с.
Потенціал, мВ
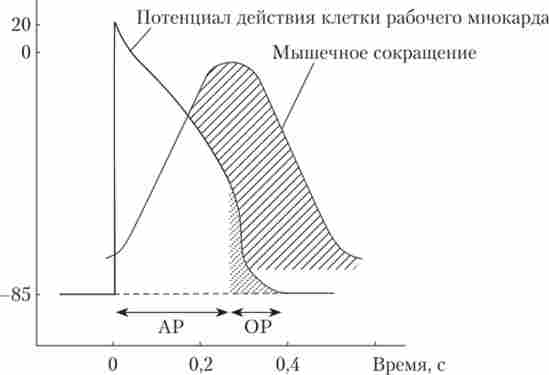
Мал. 6.12. Тимчасові співвідношення між потенціалом дії і скороченням клітини робочого міокарда шлуночка:
показані періоди абсолютної (АР) і відносної рефрактерності (ОР)
Швидкість проведення збудження по волокнах робочого міокарда відносно невелика. І в передсердях, і в шлуночках вона становить 0,3-0,5 м / с. Це в 10 разів повільніше, ніж проведення збудження по волокнах скелетного м'яза, і в 250 разів повільніше, ніж проведення по нервовому волокну. Синхронізація скорочень волокон міокарда забезпечується розвитком провідної системи.
Скорочення серцевого м'яза відрізняється від скоротливості скелетних, як за основними характеристиками, так і за механізмом запуску скорочення. Волокна серцевого м'яза коротшають на дуже тривалий час: в шлуночках - на 300-400 мс, в передсердях - на 100 мс. Час скорочення одиночного миокардиального волокна майже збігається з тривалістю ПД і рефрактерного періоду (див. Рис. 6.12). Це робить серце нездатним до тетанічних скорочення навіть при частих подразненнях, збільшуючи надійність виконання ним насосної функції.
Електромеханічне сполучення в серцевому м'язі, завдяки якому ПД призводить до скорочення міофібрил, має ряд важливих особливостей. Так само, як і в скелетних м'язах, ПД, поширюючись по клітинній мембрані, потрапляє всередину серцевих волокон по Г-трубочок (каналам Г-системи). Порушення мембрани в області Г-трубочок призводить до відкриття Са2+-каналів і надходженню в клітину серцевого м'яза іонів Са2+ з позаклітинного простору (в скелетному м'язі такий механізм відсутній). Надійшли в цитоплазму іони Са2+ взаємодіють з особливими рецепторами на мембранах саркоплазматичного ретикулума, що призводить до викиду з них власного (внутрішньоклітинного) кальцію. За тисячні частки секунди іони Са2+ досягають міофібрил і ініціюють скорочення. Таким чином, для скорочення волокон серцевого м'яза використовується не тільки внутрішньоклітинний, а й зовнішній кальцій.
Поряд з механізмом надходження іонів Са2+ із зовнішнього середовища, в кардіоміоцитах існує і ефективний механізм його видалення. При розслабленні м'язи активується робота Са2+-насоса не тільки в мембранах саркоплазматичного ретикулума, але і в зовнішній мембрані, а також Иа+/ Са2+-обмінника, який відкачує з клітки іони Са2+ за рахунок енергії що входять в клітину за градієнтом концентрації іонів Na+. Схема, що описує механізм електромеханічного сполучення в серцевому м'язі, наведена на рис. 6.13.
Будучи функціональним сінцітіем, серцевий м'яз відповідає на роздратування порогової сили максимальної реакцією, т. Е., Як і скелетне м'язове волокно, підкоряється закону «все або нічого».
Механічні властивості серцевого м'яза сприяють її успішному функціонуванню. Еластичність і розтяжність міокарда пом'якшують гідродинамічний удар крові об стінки шлуночків при їх наповненні. Розтягування стінок шлуночків надійшла кров'ю викликає виникнення еластичної тяги, що підсилює подальше скорочення. Скорочення міокарда викликає виникнення сили, яка прагне відновити звичайну довжину його волокон. Це прискорює розслаблення міокарда і приплив крові в шлуночки.
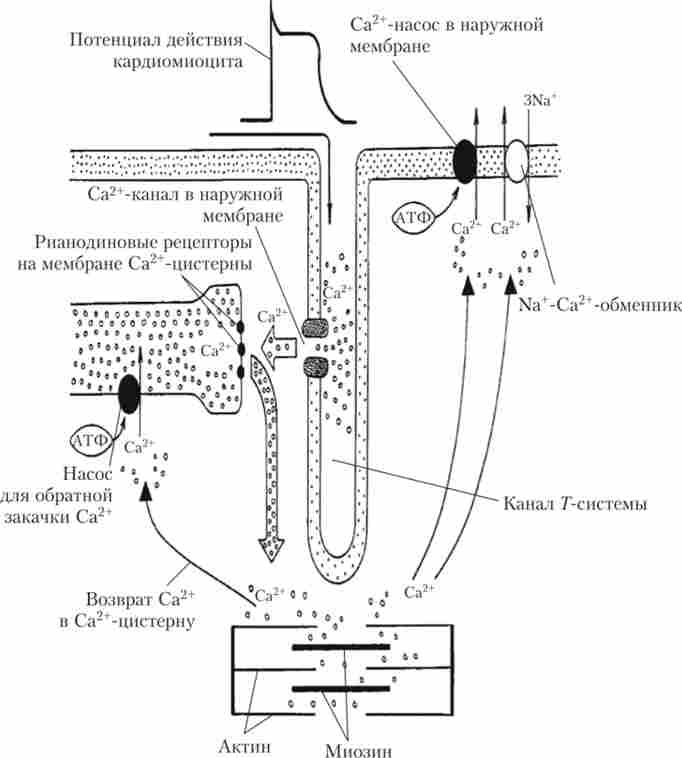
Мал. 6.13. Схема механізму електромеханічного сполучення в клітинах серцевого м'яза
серцевий цикл являє собою послідовну зміну станів скорочення (систола) і розслаблення (діастола) камер серця. Цикл починається з систоли передсердь, яка триває 0,1 с і змінюється диастолой. Одночасно з диастолой передсердь розвивається систола шлуночків (0,33 с), яка змінюється фазою загального розслаблення серця (близько 0,4 с). В цей час і передсердя, і шлуночки перебувають у розслабленому стані. Систола шлуночків складається з періоду напруги і періоду вигнання. Під час періоду напруги (0,08 с) стінки шлуночків изометрично (без зміни довжини) скорочуються, підвищуючи тиск заповнює їх крові. Вихід крові з шлуночків не допускають закриті стулчасті і півмісяцеві клапани. Тиск в лівому шлуночку досягає 60-80 мм рт. ст., а в правому - 15-20 мм рт. ст. Це дорівнює тиску в аорті і легеневої артерії, відповідно, і призводить до відкриття полумісячну клапанів.
З початком виходу крові верб серця починається період вигнання (0,25 с), який включає швидку фазу (0,12 с) і повільну фазу (0,13 с) і завершується з початком діастоли.
Діастолу шлуночків ділять на три етапи: розслаблення, наповнення і додаткове наповнення. Під час розслаблення (0,12 с) починає знижуватися напруга міокарда, а потім послідовно закриваються півмісяцеві клапани, падає тиск в шлуночках і відкриваються стулчасті клапани. З цього моменту починається наповнення шлуночків кров'ю з нескорочених передсердь, яке триває близько 0,25 с. Після вирівнювання тисків в шлуночки надходить додаткова кількість крові за рахунок систоли передсердь. Цей етап діастоли триває 0,1 с. В результаті один серцевий цикл завершується і його змінює інший. Частота зміни циклів і кількість крові, що переміщується за один цикл, є найважливішими кількісними показниками роботи серця (рис. 6.14).
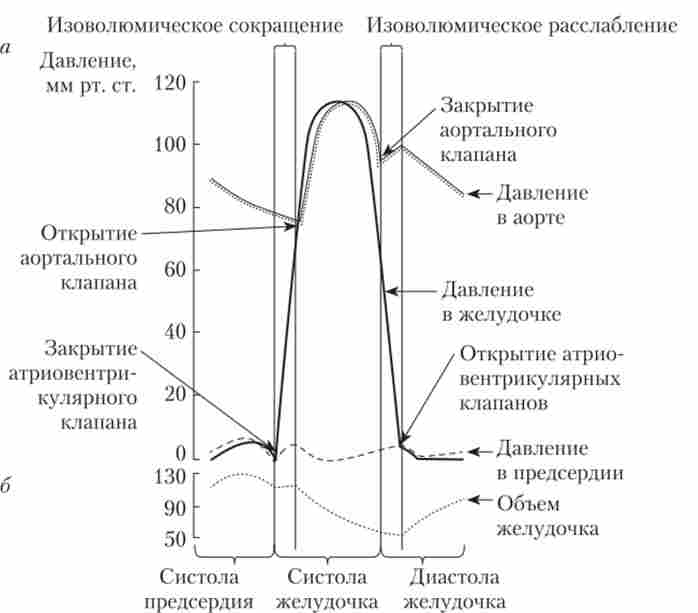
Мал. 6.14. Серцевий цикл лівого шлуночка:
а - зміна тиску в лівому передсерді, лівому шлуночку і аорті;
6 - зміна обсягу шлуночка
Кількісні показники роботи серця крім теоретичного інтересу становлять велику діагностичну цінність. Вони дозволяють оцінити ступінь тренованості серця, його адаптивні можливості, а також виявити деякі патології.
Одним з найважливіших показників є частота серцевих скорочень. У здорових дорослих людей вона складає від 60 до 80 уд / хв, у молодих - близько 70-90 уд / хв. У високотренірованних спортсменів нижня межа норми опускається до 45 уд / хв. Зниження частоти серцевих скорочень нижче 60 уд / хв називається брадикардией, підвищення частоти вище 90 уд / хв - тахікардією. Абсолютне значення і варіабельність частоти серцевих скорочень відображають баланс впливів симпатичної і парасимпатичної систем, рівень фізичного та емоційного навантаження.
Іншою важливою характеристикою серця є ударний обсяг - кількість крові, яка викидається шлуночком за одне скорочення. Ударні обсяги правого і лівого шлуночків в нормі рівні і в середньому складають 55-80 мл. При фізичному навантаженні ударний обсяг може досягати 120 мл у нетренованого людини і 200 мл - у спортсмена. Загальна кількість крові, виганяє лівим шлуночком за одну хвилину роботи, називається хвилинним об'ємом кровотоку. У нормі в спокої він дорівнює 4-6 л, а при фізичному навантаженні він може вирости до 30 л.
Електрокардіографія є метод оцінки поширення хвилі електричного збудження в серці. Вона заснована на вимірі різниці потенціалів між точками на поверхні тіла. Зареєстровану за цією методикою криву називають електрокардіограмою (ЕКГ), а прилад, за допомогою якого ведеться запис, - електрокардіографом. Є багато способів реєстрації ЕКГ. Стандартними або класичними вважаються відведення, запропоновані Виллемом Ейнтхове- ном в 1908 р При цьому датчики розташовуються в наступних стандартних положеннях: при першому відведенні - на правій руці і лівій руці; при другому відведенні - на правій руці і лівій нозі; при третьому відведенні - на лівій руці і лівій нозі (рис. 6.15).
Електрокардіографічна крива має три зубці: Р, QRS і Г, які розділені інтервалами. зубець Р відображає процес збудження передсердь. інтервал PR відповідає проходженню збудження через атріовентрикулярний вузол і становить 0,12-0,20 с. Подовження цього інтервалу вказує на патологію атріовентрикулярного вузла. Тривалість розвитку комплексу QRS зазвичай дорівнює 0,06-0,1 с і відображає проходження збудження по провідній системі шлуночків. Аномально подовжений комплекс QRS вказує на блокаду ніжок пучка Гіса. інтервал ST відповідає часу, коли весь міокард шлуночків збуджений. При цьому зазвичай пишеться нульова (ізоелектрична) лінія. Відхилення від неї свідчить про порушення збудливості однієї з ділянок міокарда шлуночків. Це відбувається, наприклад, при інфарктному пошкодженні серцевого м'яза. 7-зубець реєструється тоді, коли електричне збудження шлуночків поступово спадає. Однак саме цього моменту відповідає максимум їх механічного скорочення. На рис. 6.16 показані послідовність збудження різних ділянок міокарда в ході серцевого циклу і відображення цього процесу на електрокардіографічної кривої. Детальна розшифровка ЕКГ за допомогою комп'ютерних програм дозволяє зробити більш докладні діагностичні висновки.
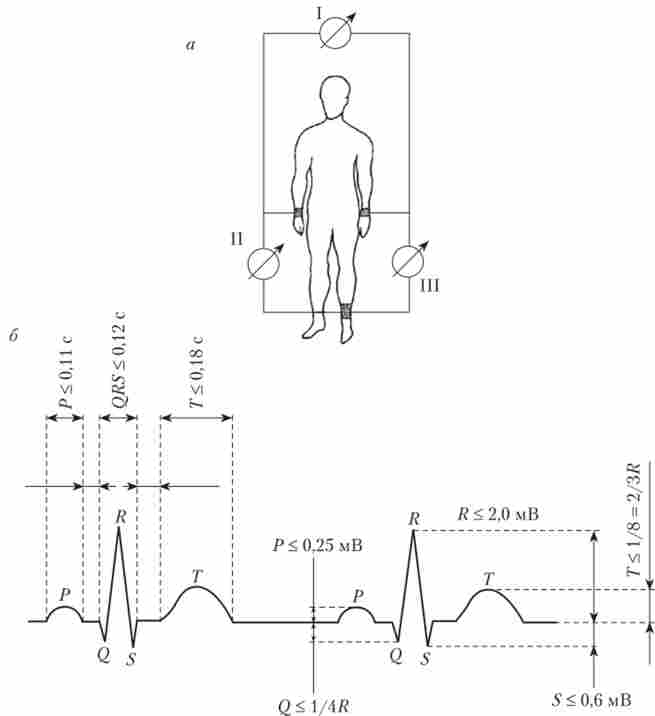
Мал. 6.15. Розташування електродів при відведенні електрокардіограми по Ейнтховеном (я); величина інтервалів і піків ЕКГ у здорової людини (II відведення по Ейнтховеном) (Б):
римськими цифрами позначені стандартні відведення
Кровопостачання серця забезпечує система коронарних судин. Коронарні артерії відходять від основи аорти. Ліва коронарна артерія постачає кров'ю ліве передсердя і лівий шлуночок, права коронарна артерія - праве передсердя і правий шлуночок, а також частково задню стінку лівого шлуночка (рис. 6.17).
Міжшлуночкової перегородки забезпечується кров'ю, що надходить і по лівій, і по правій коронарних артерій. Між гілками артерій є що з'єднують їх перемички - артеріальні анастомози (див. Рис. 6.17). Завдяки анастомозам кровопостачання міокарда не порушується навіть при пошкодженні (наприклад, тромбування) однієї з гілок
коронарних артерій. Капіляри, пов'язані з пошкодженої артерією, отримують кров через анастомози з сусідньої артерії. Таким чином, артеріальні анастомози підвищують надійність системи кровопостачання серця.
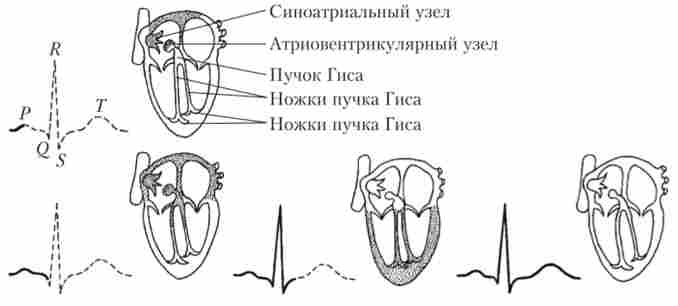
Мал. 6.16. Послідовність збудження різних ділянок міокарда і відображення цього процесу на електрокардіографічної кривої
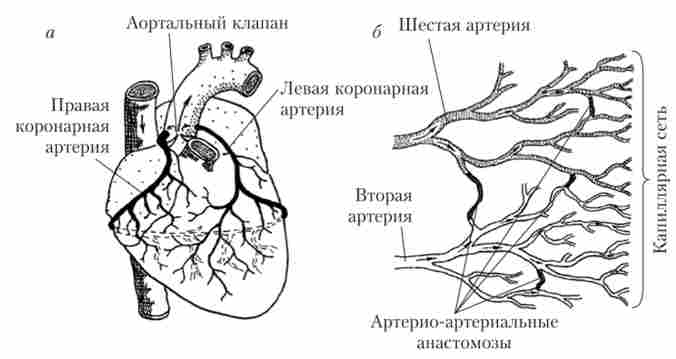
Мал. 6.17. коронарні артерії (А) і анастомози між гілками двох дрібних артерій в міокарді серця (б)
Венозна кров відтікає від серця через систему вен, які переходять в венозний синус і передні коронарні вени.
Капілярна мережа міокарда в 3-4 рази щільніше, ніж у скелетному м'язі. При масі серця, що становить 0,5% від маси тіла, воно споживає 5% крові. Коронарний кровотік сповільнюється під час систоли, коли судини пережимаються скорочується міокардом, і прискорюється під час діастоли. Це є особливо актуальним в міокарді лівого шлуночка. Скорочення діастоли, наприклад, при тахікардії створить небезпеку пошкодження міокарда.