| Головна |
| «« | ЗМІСТ | »» |
|---|
Взаємодія гормону з кліткою-мішенню починається з того, що гормон повинен сформувати гормон-рецепторний комплекс, т. е. зв'язатися з рецептором цієї клітини. Клітини, позбавлені рецепторів до якогось гормону, не здатні реагувати на гормональний вплив. Взаємодія гормону і рецептора ініціює початок каскаду ферментативних реакцій, кожна стадія якого стає все більш потужною, тому навіть мала концентрація гормону здатна давати суттєвий ефект.
Рецептори гормонів є великими білками, і кожна чутлива до гормону клітина може мати від 2000 до 100000 таких рецепторів. Рецептори високоспецифічні для конкретного гормону, що зумовлює тип гормону, який зможе вплинути на певну тканину.
Переважна кількість рецепторів до гормонів локалізується в такий спосіб:
мембранні рецептори підрозділяють на володіють власною активністю ферментативної (рецепторні тирозинкінази і гуанілат- циклази) і на що не володіють власною активністю ферментативної (пов'язані з G-білками).
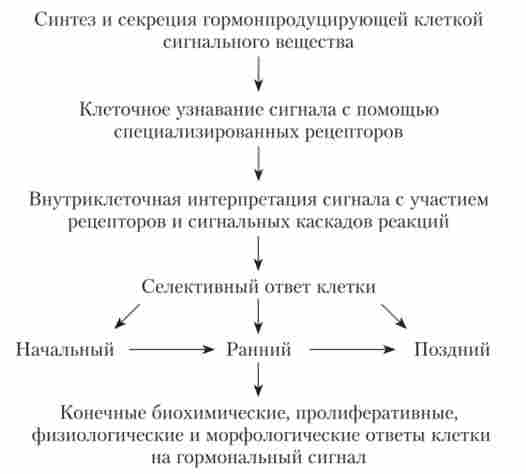
Послідовність подій, необхідних для розвитку гормонального ефекту, представлена на рис. 1.9.
Мал. 1.9. Етапи розвитку гормональних ефектів
Ядерні рецептори, які регулюють синтез білка, характерні, наприклад, для статевих гормонів (андрогени, естрогени), гормонів щитовидної залози (тиреоїдит), глюкокортикоїдів і мінералокортикоїдів наднирників. Ці гормони легко проникають через плазматичну мембрану в клітину, де в цитоплазмі зв'язуються з рецепторним білком і транспортуються в ядро. У неактивному стані підродина глюкокортикоїдних, протестеронових, естрогенових і андрогенового рецепторів буває пов'язано з білком теплового шоку (англ, heat-shock protein) HSP-90.
При зв'язуванні гормону з рецептором білок теплового шоку відділяється від останнього, комплекс білок - рецептор зв'язується з ДНК в специфічних ділянках, які називаються елементами гормонального відповіді, які розташовуються у напрямку транскрипції за сайтами початку транскрипції. В результаті транскрипція і синтез білка порушуються. Рецептори тиреоїдних гормонів і ретиноєвої кислоти в неактивному стані не пов'язані з білком теплового шоку і можуть приєднуватися до своїх елементів гормонального відповіді на молекулі ДНК під час відсутності гормонів, пригнічуючи транскрипцію. Активація рецепторів, що реалізують дію гормонів, мабуть, відбувається в результаті фосфорилювання (рис. 1.10).
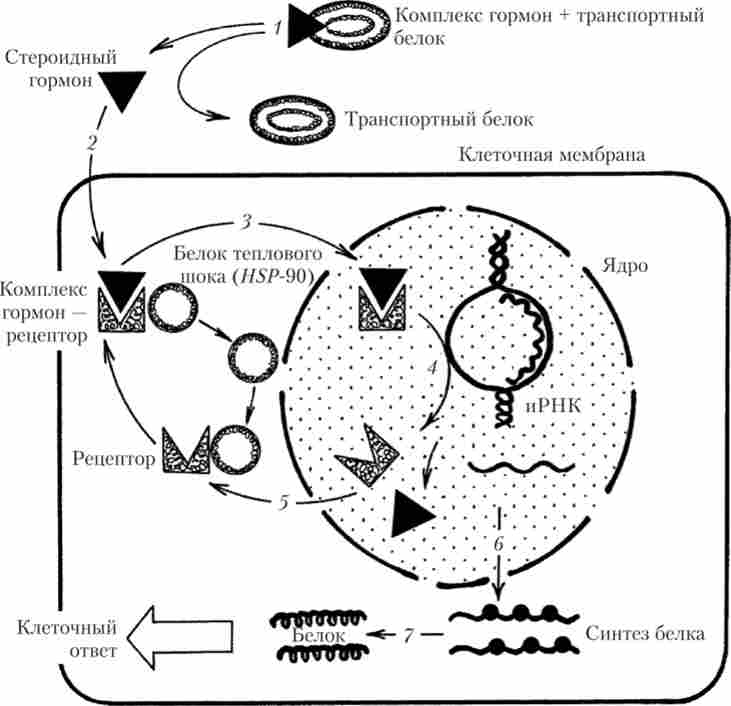
Мал. 1.10. Схема механізму реалізації ефектів стероїдних гормонів:
Гормони, не проникають через клітинну мембрану, взаємодіють з рецепторами, розташованими всередині цієї мембрани або на її поверхні.
Деякі гормони так само, як і багато медіатори, здатні активувати іонні мембранні канали, пов'язані з рецепторами або
є їх частиною (іонотропние рецептори). Взаємодія молекули такого гормону з рецептором призводить до відкриття або, рідше, закриття іонних каналів. Через відкриті канали в клітку проникають іони, найчастіше Са2+ і (або) Na+, деполярізуя клітку. Іони кальцію є найпоширенішими вторинними посередниками і потужними регуляторами внутрішньоклітинних процесів. Все це призводить до зміни стану клітини. Однак значне число гормонів, для яких характерна мембранна рецепція, які не впливають на стан іонних каналів безпосередньо, а взаємодіють з рецепторами, сполученими з G-білками або, рідше, з фермент-сполученими рецепторами (метаботропние рецептори). Схема сигналізації за допомогою іонотропних і метаботропних рецепторів приведена на рис. 1.11.
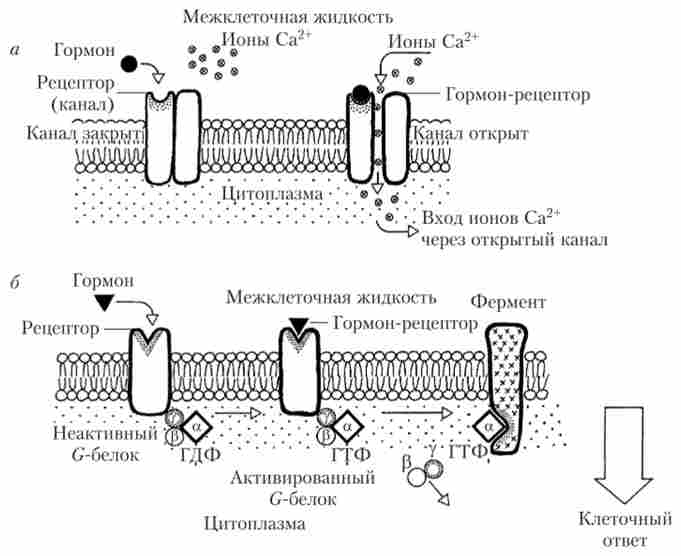
Мал. 1.11. Схема роботи гормональних рецепторів:
а - іонотропного (канатиют); б - метаботропних
Рецептори, пов'язані з G-білком, є амінокислотну послідовність, яка сім разів «прошиває» клітинну мембрану (утворюючи сім трансмембранних спіраль ділянок - доменів). Позаклітинна частина цих білків глікозильованого (рис. 1.12).
Сім трансмембранних доменів розташовані в мембрані по колу, утворюючи в центрі поглиблення зі спеціальними ділянками зв'язування ліганда. Приєднання лиганда (в нашому випадку - гормону) або агоніста призводить до зміни конформації рецепторних білків, завдяки чому активується G-білок (гуанозинтрифосфат-зв'язуючий білок). G-білок розташований на внутрішній стороні мембрани і складається з трьох субодиниць: а, Р і у. Різні G-білки розрізняються будовою а-субодиниць (див. Рис. 1.12).
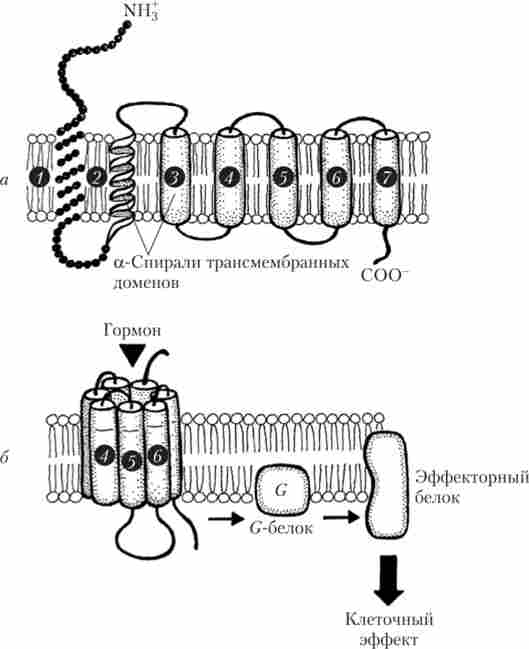
Мал. 1.12. Будова трансмембранних доменів рецепторів (1-7), пов'язаних з G-білками (а), і послідовність подій при його гормональної активації (Б)
Після приєднання гормону до рецептора конформація рецепторного білка змінюється таким чином, що а-субодиниця G-білка звільняє ГДФ (гуанозиндифосфат), приєднує ГТФ (гуанозинтрифосфат) і відділяється від Ру-субодиниць, вступаючи в контакт з еффекторним білком, змінюючи його активність. Ру-Субодиниці також можуть надавати власні ефекти. а-Субодиниця забезпечує повільний гідроліз пов'язаного ГТФ до ГДФ. комплекс СЦ-ГДФ не має спорідненості до еффекторним білків і знову возз'єднується з Ру-субодиницями. а-Субодиниці G-білків розрізняються по спорідненості і типу впливу на еффектор-
ні білки. Наприклад, Са-ГТФ Gj-білка стимулює АЦ, в той час як 6 'Ц-ГТФ З;-білка її пригнічує. За-ГТФ G ^ -белка активує фермент фосфоліпазу С (рис. 1.13).
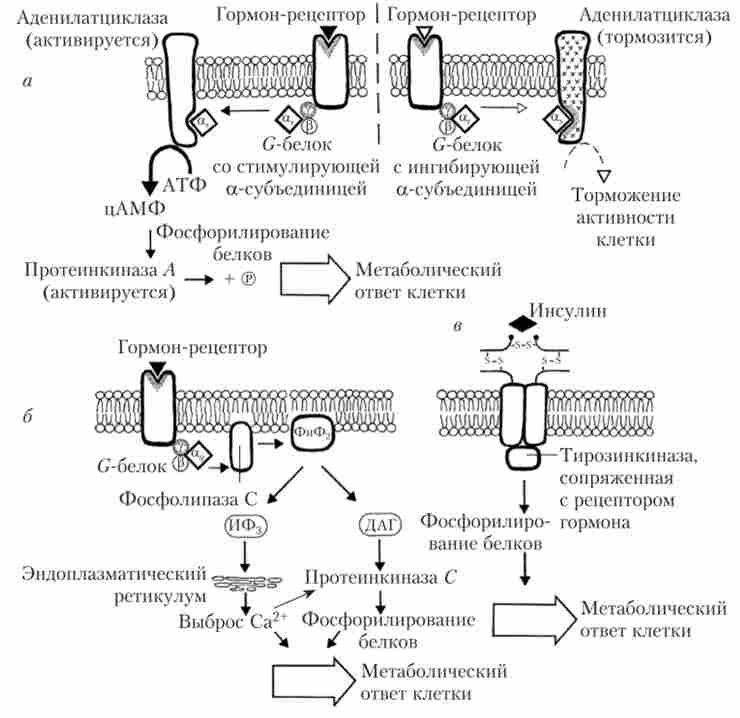
Мал. 1.13. Різні типи метаботропних рецепторів:
а - ефект опосередкований через зміну активності АЦ; б - ефект опосередковується через активацію фосфоінозитидного системи; в - ефект опосередковується через активацію тирозинкінази
G-білок-пов'язані рецептори дуже широко представлені в організмі. До них відносяться рецептори до білково-пептидних гормонів, простаглан динам, лейкотрієнів, НА, дофаміну, опіоїдів, гістаміну і багатьом іншим медіаторів і гормонів. Слід ще раз підкреслити, що ці білки відрізняються один від одного тільки будовою а-субодиниці, а в кінцевому рахунку - впливом на різні внутрішньоклітинні регуляторні системи.
У тих випадках, коли G-білок містить а ^ -субодиниці, ця субодиниця активує АЦ (рис. 1.13, а). Підвищення рівня цАМФ різко підвищує рівень життєдіяльності клітини, так як цАМФ активує протеїн Ay фосфорилюються багато регуляторні білки. Зокрема, фосфорилювання білків Са2+-каналів сприяє збільшенню надходження Са2+ до клітки. В якості вторинного посередника систему АЦ - цАМФ використовують такі гормони:
У разі деяких інших гормонів комплекс гормон - рецептор - Gq-білок активує мембранний фермент фосфоліпазу С, яка каталізує гідроліз фосфатіділікозітолдіфосфата (ФІФ2) На два важливих метаболіти, що володіють біологічною активністю: інозітолтріфосфат (ІФ3) І диацилглицерол (ДАГ) (рис. 1.13, б). ІФ3 стимулює вихід іонів кальцію з внутрішньоклітинних депо через ІФ3-рецептори, а ДАГ активація протеїн С, яка фосфорилирует цілий ряд білків, змінюючи їх активність. В цілому, спостерігаються вивільнення гормонів, ріст клітин, експресія генів і т. П. Регуляторний ефект доповнюється ще й тим, що ліпідна частина ДАГ - арахідонової кислоти - є попередником простагландинів, т. Е. Гістогормонов, що володіють багатьма регуляторними ефектами, як на місцевому тканинному, так і на системному рівнях.
У деяких клітинах G-білки можуть прямо або опосередковано впливати на молекули іонних каналів, регулюючи струм Са2+ через мембрану. Потрапивши в клітку, іони кальцію зв'язуються з білком кальмодулі- ном. Коли кальмодулин взаємодіє з чотирма іонами кальцію, його конформація змінюється, що призводить до активації або пригнічення протеинкиназ, запускаючи потужний клітинний відповідь на вплив гормону. Наприклад, міозінкіназа діє безпосередньо на міозин гладких м'язів, викликаючи їх скорочення.
Ціле сімейство рецепторів до гормонів містить вбудовані проте- інкііази, які специфічно стимулюють фосфорилирование залишків тирозину в білках-мішенях (рис. 1.13, в). До таких гормонів відносяться інсулін, інсуліноподібний фактор росту (ІФР-I) і деякі інші гормони.
Ще раз необхідно зазначити, що характер впливу гормону на клітину визначається рецепторами цієї клітини. Наприклад, при цукровому діабеті тільки близько 30% хворих відчувають нестачу інсуліну, а у решти розвивається резистентність рецепторів клітин-мішеней до цього гормону. При синдромі тестикулярной фемінізації генотип у людини відповідає чоловічому (XY), Проте фенотипично розвивається жінка. У хворих відбувається мутація двох генів, в яких закладена структура рецепторів до чоловічого статевого гормону - дигидротестостерону (ДГТ). Цим визначається неможливість для гормону, рівень якого нормальний, надати програмує ефект, і стать ембріона залишається базовим, т. Е. Жіночим. У табл. 1.4 наведені основні типи рецепторів до гормонів і приклади гормонів, що взаємодіють з цими рецепторами.
Таблиця 1.4
Основні суперсімейства гормональних рецепторів і взаємодіють з ними гормони
|
Тип рецептора |
пов'язується гормон |
|
ядерні |
Стероїдні гормони:
|
|
Тиреоїдні гормони:
|
|
|
Мембранні, пов'язані з G-білками |
|
|
Мембранні, пов'язані з тирозинкіназ |
|
|
Мембранні, рецепторні тіро- зінкіпази і рецепторні гуані- латціклази |
|