| Головна |
| «« | ЗМІСТ | »» |
|---|
У 50-х рр. XX століття Р. Емерсон припустив, що залежна від світла фаза фотосинтезу містить дві окремі фотосистеми, причому обидві вони повинні активуватися для досягнення максимальної ефективності світлових реакцій. Фотосистема I (ФС I) містить в основному хлорофіл а, поглинає при 700 нм, а фотосистема II (ФС II) - при 680-683 нм. Багато деталей світлових реакцій невідомі. Встановлено, що обидві фотосистеми виконують окремі, але взаємодоповнюючі функції. Поглинання кванта світла ФС I переводить хлорофіл-700 в електронно-збуджений стан, в ньому відбувається поділ зарядів за рахунок присутності поруч з порушеною хлорофілом допоміжних пар: окислювач-відновник. Оскільки їх природа невідома, відновлений акцептор на рис. 16.2 позначений А ^. При цьому збуджений хлорофіл перетворюється в катіон-радикал, т. З. в його молекулі утворюється «електронна дірка» за схемою:
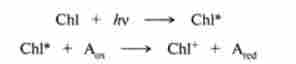
де Chi, Chi *, Chl+ - молекули хлорофілу в основному, електрон но-збудженому і катіон-радикальному станах. Високоенергетичні електрони з Ared, утвореного в ФС1, відновлюють железосерний негемове білок ферредоксин, який бере участь у відновленні НАДФ * таким чином, що НАДФ Н відразу надходить в строму. В реакції відновлення НАДФ 'ферредоксин (FD) бере участь ФАД-залежна дегидрогеназа - ферредоксин: НАДФ'-оксидоредуктаза:

У фотосистемі II за збудженням молекули хлорофілу слід передача порушеної електрона на пару окислювач-відновник реакційного центру ФСП. При цьому утворюється катіон-радикал хлорофілу і сильний
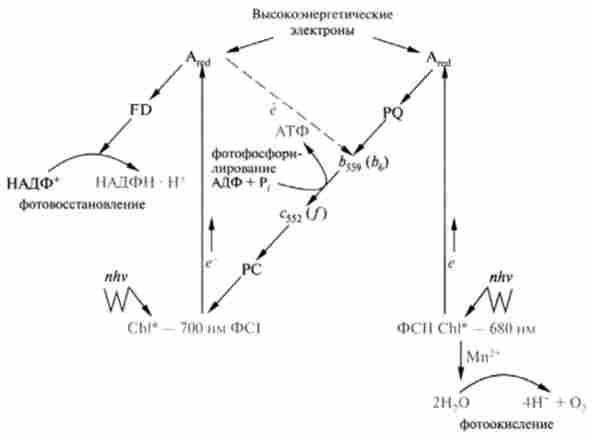
Мал. 16.2. Z-Схема потоку електронів в моделі двох фотосистем (ФС 1 і ФС II):
FD - феррелоксін; PQ - пластохинон: PC - пластостоііаніі; 6559(66) - і з552(/) - цитохроми; Chi * - електронно-збуджений хлорофіл; Ared - відновлений акцептор електронів; пунктиром показаний механізм циклічного фосфорилювання
окислювач, який бере участь в окисленні молекули Н20 до 02, а електрони води відновлюють Аох до Ared і далі надходять в ланцюг переносників електронів від ФС II до ФС I. Важливу роль в окисленні Н20 в реакційному центрі ФС II грають іони марганцю, виконуючи, мабуть, роль пари окислювач / відновник.
Найближчим акцептором електронів, генерованих ФСП, є пластохинон (PQ) - структурний аналог убихинона (KoQ):
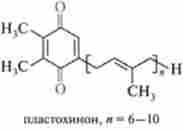
Потім в ланцюзі перенесення електронів беруть участь два цитохрому Ь559(Ьв) і з552(/); наступним переносником електронів є пластоціанін (PC) - медьсодержащий білок, що передає електрон в реакційний центр ФС I і заповнює в ньому «електронну дірку». Частина енергії збуджених електронів, що проходять через комплекс, утворений PQ, цитохромами />559, з552 і PC, трансформується в хімічну енергію АТФ, т. е. йде процес фотофосфорилування.
Механізм фотосінтстічного фосфорилювання схожий з синтезом АТФ в процесі окисного фосфорилювання в мітохондріях. Система переносників електронів інтегрована в мембрану тилакоида таким чином, що перенесення пари електронів створює потік протонів з зовнішньої поверхні тилакоида всередину, pH на внутрішній поверхні тилакоида може досягати 4 і нижче. Таким чином, на мембрані створюється електрохімічний протонний потенціал ДРН *, який використовується інтегрованої в мембрану Незалежної синтетазой для синтезу АТФ (рис. 16.3). Структура цього ферменту аналогічна мітохондріальної АТФ-синтетази (гл. 15) і зазвичай позначається як CF0-CF ,. Символ З означає, що цей ферментний комплекс локалізована в хлоропластах (chloroplast) І, подібно до мітохондріальної Н+-залежною-АТФ-синтетази, включає гідрофобний, інтегрований в мембрану тилакоида компонент (CF0) І гідрофільний комплекс (CF,), що каталізує синтез АТФ.
Як видно з рис. 16.3, електрони рухаються до зовнішньої сторони мембрани, а протони концентруються на внутрішній поверхні тилакоїдів, т. Е. Напрям протонного градієнта ДрЬг протилежно напрямку його в мітохондріях. Таким чином, тилакоїди представляють собою як би вивернуті навиворіт мітохондрії, тому АТФ утворюється з їх зовнішнього боку і безперешкодно надходить в строму для участі в темнових стадіях фотосинтезу.
Синтез АТФ, пов'язаний з функціонуванням обох фотосистем, отримав назву нециклічного фосфорилювання. Слід пам'ятати, що заповнення дефіциту електронів в ФС I відбувається за рахунок перенесення до неї електронів води, що надходять від ФС II. Процес нециклічного фосфорилювання пов'язаний з фотовідновлення НАДФ+.
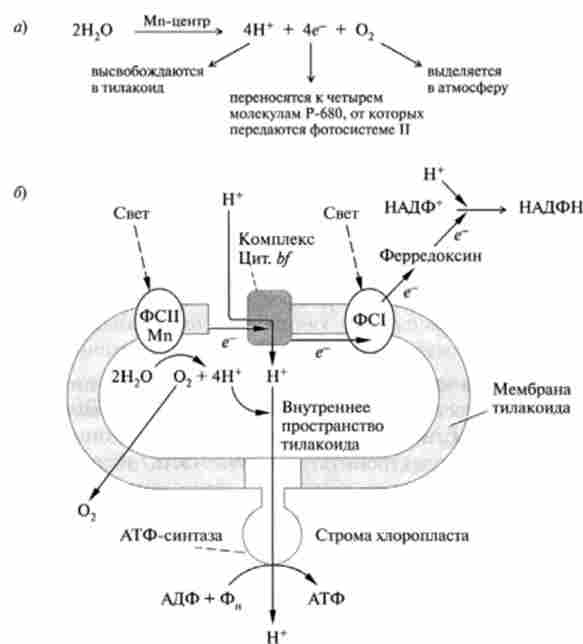
Мал. 16.3. Схема процесів, що відбуваються в Тилакоїди: а - сумарна реакція в Мп2+-містить центрі; 6 - доля протонів і електронів
(По В. Елліот, Д. Еліот)
Однак функціонує додатковий механізм, що забезпечує синтез АТФ без супутнього відновлення НАДФ+, в якому бере участь тільки ФС I (див. рис. 16.2). Потік електронів, що надійшли з реакційного центру ФС I і відновили А ^, повертається в ФС I, проходячи через цитохром и bSS9 і /, пластоціанін, і, створюючи протонний градієнт на тілакоідной мембрані, використовується для синтезу АТФ. Оскільки в даній системі спостерігається циклічний потік електронів, то цей шлях синтезу АТФ отримав назву ціюшчного фотосинтетичного фосфорилювання. Отже, цей шлях синтезу АТФ не сполучений з відновленням НАДФ виділенням 02 і відбувається в тому випадку, коли клітина забезпечена НАДФН, але відчуває потребу в АТФ.