| Головна |
| «« | ЗМІСТ |
|---|
Клітини різних організмів, що живуть на Землі, і навіть одного і того ж організму можуть дуже сильно відрізнятися і за розмірами, і але будовою, і але функціям. Наприклад, порівняйте нейрон, клітку серцевого м'яза, еритроцит і сперматозоїд (рис. 1.13).
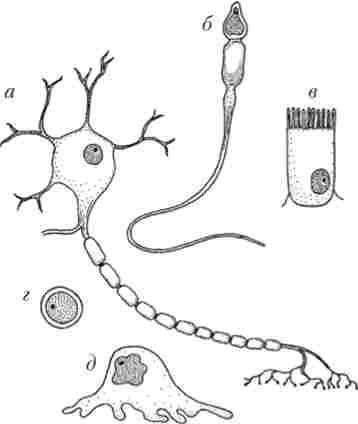
Мал. 1.13. Приклади клітин різної форми:
а - нейрон; 6 - сперматозоїд; в - ентероціт; г - лейкоцит; д - фагоцит
Однак подібностей між клітинами більше, ніж відмінностей.
Всі клітини поділяють на еукаріотичні, мають ядро, і прокариотичні, оформленого ядра не мають. Оскільки всі клітини організму людини, за винятком зрілих еритроцитів, мають ядро, в подальшому буде охарактеризована еукаріотична клітина.
У внутрішній полужидкой середовищі клітини - гіалоплазме - присутній цілий набір структур, що виконують функції, необхідні для забезпечення нормальної життєдіяльності клітини: харчування, дихання, виділення продуктів обміну, розкладання, якщо треба - руху. Для цього всі клітинні структури необхідно утримувати разом, щоб клітина могла існувати як єдине ціле. Це завдання вирішує клітинна мембрана, що покриває клітку.
Зовнішня клітинна мембрана (плазматична мембрана, плаз- малемма), товщиною 8-12 нм, має подібну будову у всіх клітин і побудована з двох шарів фосфоліпідів. Кожна молекула фосфолипида в своїй структурі має гідрофільну головку і гідрофобний хвіст. У біологічних мембранах молекули ліпідів розташовуються головками назовні, а хвостами всередину. Численні молекули білків занурені в біліпідний шар мембрани (рис. 1.14).
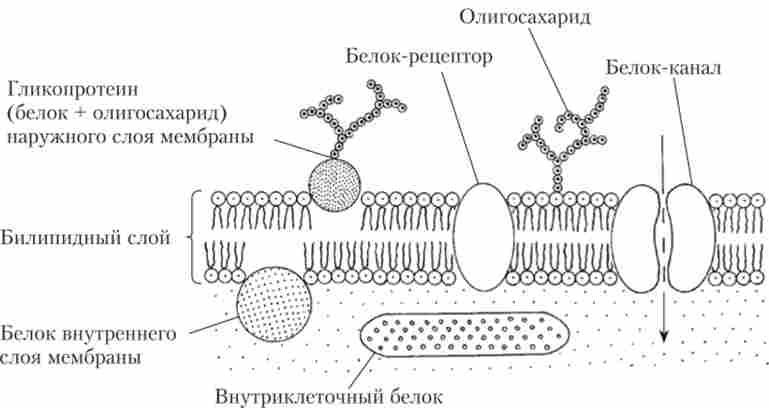
Мал. 1.14. Схема ділянки клітинної мембрани (основні типи мембранних білків)
Одні білки розташовуються на зовнішній стороні мембрани, інші прилягають до внутрішньої, а треті пронизують мембрану наскрізь. Частина мембранних білків є рецепторні структури, за допомогою яких клітина сприймає різні сигнальні впливу, інші утворюють трансмембранні канали, за якими йде пересування різних іонів в клітину і з неї (іонний струм). Третя група білків - це ферменти, що забезпечують протікання процесів життєдіяльності клітини.
До складу мембран входить невелика кількість вуглеводів (близько 5%). Це зазвичай глікопротеїди і гліколіпіди, які беруть участь у виконанні зовнішньої мембраною рецепторних функцій.
Цитоплазма являє собою вміст клітини від внутрішньої поверхні клітинної мембрани до ядра і включає в себе гіалоплазму, органели (постійні структурні компоненти) і клітинні включення (тимчасові компоненти).
У цитоплазмі клітин еукаріот обов'язково є складна опорна система - цитоскелет, підтримує форму клітини і забезпечує внутрішньоклітинний транспорт. Він зібраний з трьох видів елементів: микротрубочек, проміжних філаментів і мікрофіламентів.
Микротрубочки пронизують всю цитоплазму і являють собою порожнисті трубки діаметром 20-30 нм, що складаються з спірально закручених ниток білка тубуліну. Микротрубочки утворюють основу цитоскелета і беруть участь в перенесенні але цитоплазмі різних речовин.
Проміжні філаменти мають товщину близько 10 нм, а мікрофіла- менти - 4 нм. Їх основа - білок актин. Мікрофіламенти часто зібрані в пучки і розташовуються поблизу зовнішньої клітинної мембрани. Вони здатні змінювати її форму, що дуже важливо в процесі фагоцитозу або піно- цитоза.
Центриоль є найтоншу білкову трубочку довжиною 0,3 мкм, стінки якої складаються з дев'яти груп але три мікротрубочки, з'єднаних між собою поперечними білковими сшивками. Центриоль в підставі вії або джгутики називається базальним тільцем. У багатьох клітинах дві центріолі з'єднуються по дві і стають центром організації цієї клітини, утворюючи ЦЕНТРОС (Клітинний центр) (рис. 1.15).
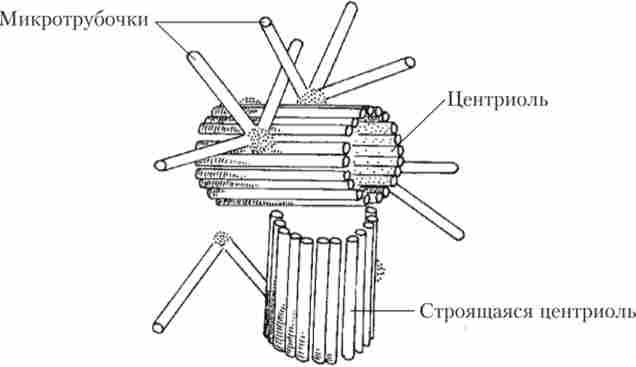
Мал. 1.15. Клітинний центр з подвоюється центріолио
Центриоли беруть участь в побудові цитоскелету і формуванні веретена поділу при утворенні нових клітин. Перед поділом клітини центріолі розходяться до полюсів клітини, і кожна з них добудовує дочірню центриоль.
Фахівці з еволюції вважають, що перші клітини, що з'явилися на Землі, мали тільки зовнішню мембрану. Їх органели ще не включали до свого складу мембран. Компоненти клітини, побудовані з мембран, з'явилися пізніше.
Ендоплазматична мережа, або ЕПР, виглядає як мережа тільки в мікроскоп. Насправді це розгалужена система сполучених порожнин і трубочок, утворених мембраною, подібна до плазматичної (рис. 1.16).
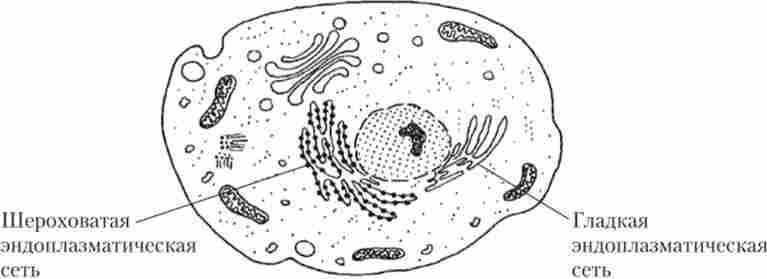
Мал. 1.16. ендоплазматична мережа
ЕПР пов'язаний із зовнішнім шаром двуслойной мембрани клітинного ядра. На частині мембран ЕПР розташовуються рибосоми, що синтезують білки, які надходять в порожнині ЕПР. Такий ЕПР називають шорстким. Чи не містить рибосом ЕПР називається гладким; він є місцем синтезу ліпідів і вуглеводів. Синтезовані в ЕПР речовини по системі трубочок транспортуються до місць використання чи накопичення.
Рибосоми представляють собою органели, що забезпечують процес синтезу білка. Їх розмір становить приблизно 20 х 30 нм. В одній клітці може міститися одночасно кілька мільйонів рибосом. Кожна рибосома складається з малої і великої субодиниць, утворених рРНК і білками. Субодиниці рибосом формуються в ядрі в області ядерець і через пори в ядерній оболонці надходять в цитоплазму. Рибосоми здійснюють збірку білкових молекул білків з амінокислот, що доставляються до рибосоми за допомогою тРНК. Між рибосомальною субодиницею є щілина, в якій розміщується мРНК, а на великий субодиниці є борозенка, по якій сповзає синтезируемая молекула білка (рис. 1.17).
Таким чином, рибосоми здійснюють трансляцію генетичної інформації - своєрідний «переклад на мову амінокислот з мови нуклеотидів».
Рибосоми або присутні в цитоплазмі у вільному стані, або розташовуються групами па мембранах ЕПР. Вільно розташовані рибосоми синтезують білки для потреб самої клітини, а прикріплені рибосоми - білки, призначені для використання у позаклітинному просторі або в інших клітинах організму.
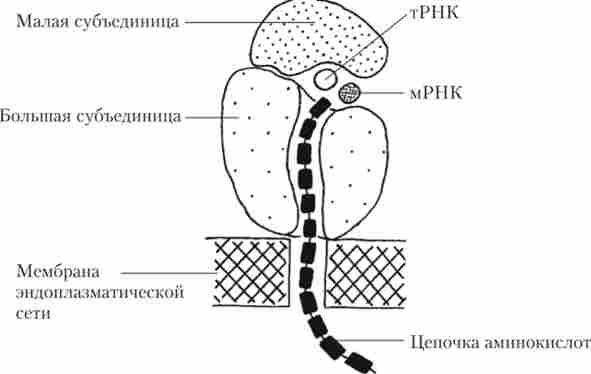
Мал. 1.17. Участь рибосоми в синтезі білка
Апарат (комплекс) Гольджі - система внутрішньоклітинних цистерн, відмежованих від цитоплазми біологічними мембранами. Комплекс Гольджі здійснює накопичення, біохімічні перетворення і упаковку в везикули (мембранні пухирці) речовин, що виробляються в клітці. Упаковані речовини переносяться в різні місця цитоплазми або транспортуються до клітинної мембрани і виводяться за межі клітини.
Комплекс Гольджі включає в себе систему цистерн (плоских порожнин), укладених стопками одна на іншу (рис. 1.18).
Мал. 1.18. Схема будови апарату Гольджі
Апарат Гольджі розташовується поруч з ЕПР, але не повідомляється з нею, тому всі речовини, синтезовані на мембранах ЕПР, переносяться
в комплекс Гольджі всередині везикул, що відділяються від ЕПР. Одна з важливих функцій апарату Гольджі - збірка клітинних мембран. Мембранні білки і ліпіди надходять в комплекс Гольджі з ЕПР, де збираються ділянки мембран, з яких виготовляються особливі мембранні бульбашки. Ці бульбашки пересуваються по цитоплазмі в ті ділянки клітини, де потрібно добудувати мембрану. Бульбашки вбудовуються в мембрану, формуючи своєрідні «латки», що ліквідовують дефекти в мембрані.
Лізосоми. Харчові частинки, які потрапляють в клітину, необхідно переварити, т. Е. Зруйнувати до таких складових частин, які клітина може використовувати. Для цього фагоцитарний пляшечку з харчової часткою повинен злитися з лизосомой. Лізосома - це маленька Везикула діаметром 0,4-1 мкм, що містить близько 50 різних видів травних ферментів, здатних розщеплювати білки, вуглеводи, ліпіди, нуклеїнові кислоти. Формуються лізосоми в комплексі Гольджі.
Відокремилися від комплексу Гольджі лізосоми, що містять ферменти в неактивній формі, називаються первинними. Везикули, що утворилися при злитті первинних лізосом з фагоцитарних вакуолями, називаються вторинними або травними вакуолями (рис. 1.19).
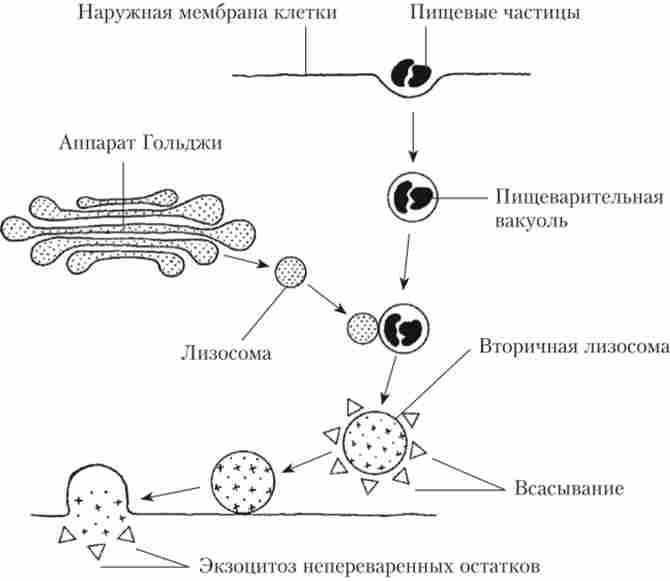
Мал. 1.19. Схема перетравлення в клітці
У них активовані ферменти здійснюють процеси перетравлення харчових речовин, які потім надходять в цитоплазму, а все, що не може бути перенесено з вторинної лізосоми в клітку, викидається назовні шляхом екзоцитозу.
Мітохондрії - це органели, в яких за участю кисню великі органічні молекули окислюються до вуглекислого газу і води. Виділяється в цих процесах енергія йде на синтез АТФ - універсального біохімічного «акумулятора» енергії. АТФ зберігається в клітці до тих пір, поки не буде потрібно. Мітохондрії зустрічаються практично у всіх клітинах еукаріот. Число мітохондрій в клітині варіює від одиниць (сперматозоїди, найпростіші і деякі водорості) до декількох тисяч (клітини печінки, м'язові клітини). Мітохондрії можуть мати кулясту, овальну або паличкоподібну форму.
Незалежно від форми, все мітохондрії мають загальний план будови. Вони утворені двома мембранами. Зовнішня мембрана гладка, а внутрішня утворює велике число складок (крист), які збільшують площу поверхні. На кристах розташовуються ферментативні комплекси, що забезпечують процеси клітинного дихання, що призводять до синтезу АТФ (рис. 1.20).
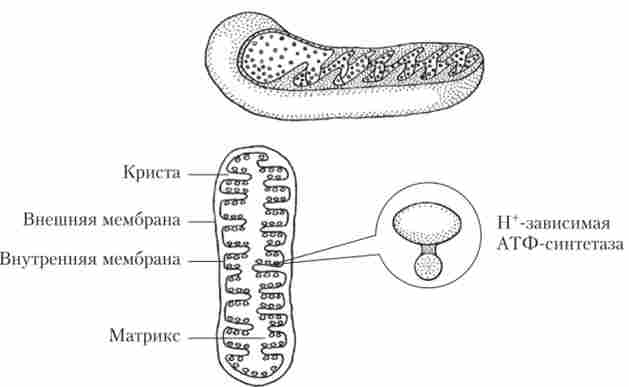
Мал. 1.20. будова мітохондрії
Мітохондрії є єдиними органеллами тваринної клітини, що мають власні ДНК, РНК і особливі рибосоми. Ця локальна генетична система забезпечує самовідтворення мітохондрій. Мітохондріальна ДНК замкнута в кільце. У періоди, коли клітці потрібна додаткова енергія, кількість мітохондрій в ній помітно зростає, а під час знижених енерговитрат - зменшується. Кільцева ДНК, здатність до самовідтворення, а також відсутність мітохондрій у прокаріотів - все це дозволило припустити, що мітохондрії походять від паразитичних прокаріотів істот, що нагадують сучасні бактерії. В процесі еволюції симбіоз змінив паразитизм, і «колишні» прокариотичні клітини перетворилися в органели, характерні для всіх еукаріотичних клітин.
Спільні риси з мітохондріями мають пластиди - органели, характерні тільки для рослинних клітин.
Органели, які здійснюють руху. Багато клітини здатні до переміщення в просторі, причому механізми рухових реакцій можуть бути різними.
Сперматозоїди тварин рухаються за допомогою джгутика довжиною близько 100 мкм. На поперечному зрізі джгутика помітно, що на периферії розташовуються дев'ять пар мікротрубочок, а в центрі - одна пара. Всі пари мікротрубочок пов'язані між собою за допомогою білка, здатного за рахунок енергії АТФ змінювати свою просторову структуру. В результаті цього процесу пари мікротрубочок переміщаються один щодо одного, згинається джгутик і сперматозоїд починає рух.
Подібний механізм забезпечує руху війок, якими забезпечені клітини багатьох органів. Довжина вій становить всього 10-15 мкм. На мембрані однієї клітини їх може бути дуже багато, але при цьому їх руху зазвичай чітко скоординовані. Так, наприклад, клітина війкового епітелію верхніх дихальних шляхів людини має близько 250 війок.
Клітинні включення формуються з речовин, які клітина або витрачає сама, або виводить у зовнішнє середовище. Прикладами клітинних включень можуть бути краплі жиру, зерна глікогену, білкові гранули і т. Д. Зазвичай вони розташовані безпосередньо в цитоплазмі і не відділені від неї мембраною. Клітинні включення використовуються органеллами клітини для харчування та інших потреб.
Ядро є найважливішою структурою еукаріотів. Воно являє собою центр управління клітини і сховище інформації про неї: в ядрі локалізовано понад 90% клітинної Д11К. При видаленні ядра клітина швидко гине. Зазвичай ядро розташовується в центрі клітини і має кулясту форму (рис. 1.21).
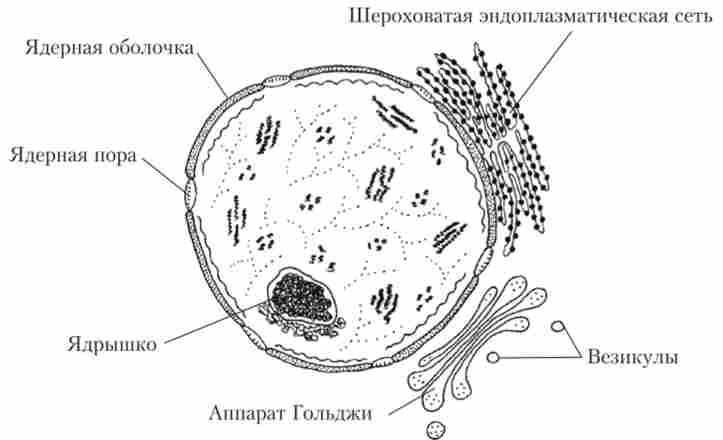
Мал. 1.21. Схема будови ядра
Діаметр ядра складає від 2 мкм до декількох десятків мікрометрів в залежності від різновиду клітини.
Ядро відокремлене від цитоплазми двумембранной оболонкою, товщина якої становить близько 30 нм. Зовнішня мембрана в складі оболонки плавно переходить в камери ЕПР і містить ядерні пори, а внутрішня - щодо гладка. Пори забезпечують вихід з ядра в цитоплазму мРНК і тРНК, а з цитоплазми в ядро через пори можуть проникати іони і більші молекули - АТФ і різноманітні ферменти.
Гелевидний вміст ядра носить назву каріоплазми. У кариоплазме розташовуються молекули ДНК, з'єднані з особливими білками - гистонами. Разом вони утворюють ядерний хроматин. ДНК людини в розгорнутому вигляді має довжину близько 5 см. Перед поділом клітини кожна молекула ДНК утворює хромосому, для чого щільно скручується і укладається за допомогою гістонів. Обсяг, займаний ДНК, при утворенні хромосом багаторазово зменшується. Під об'єктивом світлового мікроскопа під час метафази хромосоми мають вигляд довгастих паличковидних тілець, які за допомогою перетяжок (центромер) діляться на два плеча (рис. 1.22).
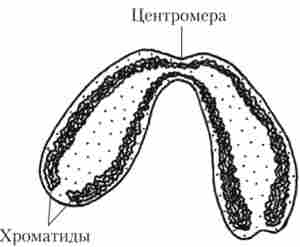
Мал. 1.22. будова хромосоми
У проміжках між поділами хромосоми розкручені (деспіралі- зовано), що забезпечує успішне функціонування генів, які представляють собою ділянки молекули ДНК, що кодують структури різноманітних білкових молекул.
Ділянки деяких молекул ДНК приймають форму петель. Такі петлі, зближуючись і перекриваючи, утворюють видимі під мікроскопом ядерця. У ядерцях синтезуються субодиниці рибосом, що потрапляють в цитоплазму через ядерні пори. У цитоплазмі формуються функціональні рибосоми, здатні брати участь в синтезі білків. Число ядерець в одній клітці може досягати семи.
Таким чином, ядро клітини виконує дві функції: