| Головна |
| «« | ЗМІСТ | »» |
|---|
Головний мозок - найбільший відділ ЦНС, розташовується в черепній коробці людини і займає 80% її обсягу. Маса його у дорослої людини коливається від 1,1 кг до 2 кг.
Усередині головного мозку розташовані порожнини - шлуночки мозку, зовні головний мозок захищений трьома мозковими оболонками, між якими знаходиться рідина. Від головного мозку відходять 12 пар черепномозкових нервів, що іннервують різні ділянки тіла. Головний мозок людини складається з п'яти відділів: довгастого, заднього, середнього, проміжного і кінцевого мозку (рис. 2.9). Два останніх іноді об'єднують назвою «передній мозок».
Довгастий мозок - нижній відділ головного мозку, що межує зі СМ. Містить численні ядра і провідні шляхи. У складі довгастого мозку знаходяться ядра слуховий, вестибулярної, смаковий, інтероцеп- тивной, шкірної і м'язової сенсорних систем, моторні ядра, що іннервують область голови, обличчя, шиї, а також вегетативні ядра, пов'язані
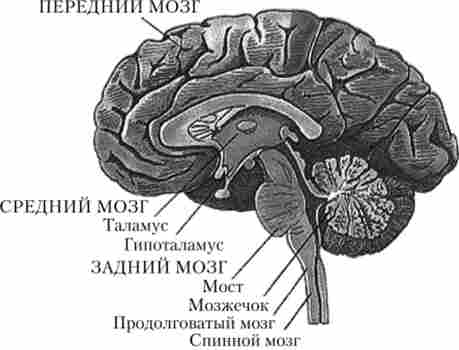
Мал. 2.9. Відділи головного мозку
з регулюванням слинних залоз, органів грудної та черевної порожнин. Довгастий мозок за рахунок специфічних нервових ядер і ретикулярної формації бере участь в реалізації вегетативних, соматичних, смакових, слухових і вестибулярних рефлексів; бере участь в реалізації ряду захисних рефлексів, таких як блювотний, чхальний, кашльовий, сльозовиділення, змикання повік.
Велика частина вегетативних рефлексів довгастого мозку реалізується через розташовані в ньому ядра блукаючого нерва, які отримують інформацію про стан діяльності серця, частини судин, травного тракту, легенів, травних залоз і ін. У відповідь на цю інформацію ядра організують рухову або секреторну реакцію названих органів. У довгастому мозку локалізується центр слиновиділення, його парасимпатична частина забезпечує посилення загальної секреції слинних залоз, а симпатична - підсилює їх білкову секрецію. Тут же знаходяться дихальний і судиноруховий центри.
дихальний центр кожної симетричною половини довгастого мозку розділений на дві частини - вдиху і видиху. Нейрони дихального центру діляться на інспіраторні (Вдих) і експіраторние (Видих). Аферентні сигнали до нейронів дихального центру йдуть від діафрагми, міжреберних м'язів, верхніх дихальних шляхів, рецепторів альвеол (через блукаючий нерв), рецепторів судин, особливо від зони біфуркації сонних артерій. Еферентні шляху з дихального центру йдуть до мотонейронам передніх рогів протилежного боку СМ, забезпечуючи скорочення діафрагми і міжреберних м'язів.
В судиноруховий центр афферентация йде від рецепторів судин, від бронхіол, серця, органів черевної порожнини, рецепторів соматичної системи. Еферентні шляху рефлексів проходять по ретікулоспі- налиюму тракту до бічних рогах СМ. Ефект зміни кров'яного тиску залежить від того, які нейрони збуджуються, і від частоти генерації імпульсів. Високочастотна імпульсація підвищує, а низькочастотна - знижує кров'яний тиск. Низькочастотна стимуляція симпатичних нейронів СМ, на яких закінчуються ретікулоспінальние шляху від сосудодвігателиюго центру, знижує тонус судин, високочастотна - підвищує його. Порушення судинного центру змінює дихальну ритміку, тонус бронхів, м'язів кишечника, сечового міхура.
Задній мозок - відділ головного мозку, розташований між середнім і довгастим. Складається з варолиева моста і мозочка.
варолиев міст - це вентральна частина заднього мозку. Міст утворює структури ростральної частини дна IV шлуночка. Дорсальная поверхню моста являє собою верхній трикутник ромбовидноїямки. Порожнина ромбовидноїямки рострально звужується і переходить в водопровід середнього мозку. Зверху порожнину ромбовидноїямки прикрита верхнім мозковим вітрилом, який разом з нижнім мозковим вітрилом і судинним сплетінням утворює дах IV шлуночка, що має вигляд шатра. Латеральні стінки IV шлуночка в області моста утворені середніми і верхніми ніжками мозочка. Від вароліевого моста відходять чотири пари черепно-мозкових нервів: V - трійчастий нерв; VI - нерв, що відводить, VII - лицевий нерв, VIII - предверние-улітковий, або слуховий, нерв. Основна функція - провідникова.
мозочок - великий відділ заднього мозку, яка формує три головні впливи на організм: 1) на руховий апарат; 2) аферентні системи; 3) вегетативну нервову систему. Роль мозочка полягає в регуляції пози, м'язового тонусу і рівноваги; сенсомоторної координації іозних і цілеспрямованих рухів; координації швидких цілеспрямованих рухів по команді з кори великих півкуль головного мозку. Медійна червячная зона мозочка пов'язана з управлінням тонусу пози і рівноваги тіла. Надходить аферентна інформація, сигналізує про позу і стан локомо апарату. Після обробки інформації з кори мозочка команди йдуть до вестибулярному ядру Дейторса, ретикулярної формації стовбура - звідти до центрів СМ по ретікулоспінальний п вестібулоспінальних трактах.
Проміжна зона кори мозочка пов'язана зі Інтернейрони ядром і має аферентні входи від спінальних трактів. Через ядра моста проміжна зона кори мозочка отримує інформацію від рухової області кори великих півкуль. Ця інформація надходить через колатералі кортикоспинального тракту і сигналізує про підготовлюваний цілеспрямованому русі. Зіставлення приходить по цих шляхах інформації дозволяє проміжній зоні мозочка брати участь в координації цілеспрямованих рухів з рефлексами підтримки пози, виборі найбільш оптимальної для виконання руху пози. Спадні команди від проміжної зони мозочка через вставні ядро йдуть до червоного ядра і далі по руброспінальний тракту до моторних центрам СМ.
Кору мозочка утворюють три шари: зовнішній - молекулярний, середній - гангліонарний і внутрішній - зернистий. З підлягає білої речовини в кору проходять аферентні лазять і моховидні волокна, а виходять аксони грушовидних клітин (табл. 2.2).
Филогенетично молодий є латеральна кора півкуль мозочка. Через церебромостомозжечковий тракт вона отримує аферентних вхід від асоціативних зон кори великих півкуль головного мозку. У кору мозочка надходить інформація про задум руху. В півкулях і зубчастому ядрі інформація перетворюється в програму руху, яка по мозжечковоталамокортікальному тракту надходить в рухові
Зв'язки нейронів в корі мозочка
|
Тип нейронів |
шар |
аферентні зв'язку |
Контакти аксонів |
|
зірчасті клітини |
молекулярний |
Паралельні волокна від зернистих клітин |
Дендрити грушовидних нейронів в поверхневих частинах молекулярного шару |
|
Корзінчатис клітини |
Псрікаріони грушовидних нейронів (через корзіічатие колатералі) |
||
|
грушоподібні нейрони |
гангліозний |
лазячі волокна |
Глибокі ядра (більшість аксонів) |
|
Розгалуження аксонів зернистих клітин |
Вестибулярні ядра довгастого мозку |
||
|
Волокна корзинчатих клітин |
Грушоподібні нейрони і зірчасті клет- ки з короткими аксонами (поворотні колатералі грушовидних нейронів) |
||
|
Аксони зірчастих клітин |
|||
|
11орадреіергічні волокна від блакитного ядра |
|||
|
Зірчасті клітини з короткими аксонами |
зернистий |
Моховидні і лазять волокна з дендритами в зернистому шарі |
Клубочок мозочка (синапси на розетках Моховидних волокон) |
|
Клітини-зерна |
Паралельні волокна з дендритами в молекулярному шарі |
||
|
Закінчення Моховидних волокон в клубочках мозочка |
Дендрити грушовидних нейронів, корзинчатих і зірчастих клітин. Дендрити зірчастих клітин з коротким і аксонами в молекулярному шарі |
зони кори великих півкуль. Руховий акт реалізується за рахунок низхідних команд, що йдуть від рухової кори в СМ по кортікоспінальних тракту. Від зубчастого ядра мозочка інформація йде прямо до СМ. Складна система регуляції рухів з наявністю зворотних зв'язків між мозочком і корою великих півкуль дозволяє півкулях мозочка брати участь в організації швидких цілеспрямованих рухів.
Мозочок включений в систему контролю вісцеральних функцій. У дослідженнях вітчизняного фізіолога, академіка Л. А. Орбелі (1949) показано, що роздратування мозочка викликає ряд вегетативних рефлексів: розширення зіниць, підвищення артеріального тиску і т. Д.
Видалення мозочка призводить до порушення серцево-судинної діяльності, дихання, моторики та секреції шлунково-кишкового тракту. У мозочку виявлені вісцеральні проекції внутрішніх органів. При частковому або повному ураженні мозочка спостерігається три типи симптомів: атонія, астенія, астазія.
Атонія проявляється в ослабленні м'язового тонусу; зазвичай супроводжується симптомом астенії, яка характеризується слабкістю і швидкою стомлюваністю м'язів. Астенія, або синдром хронічної втоми, - хворобливий стан, що виявляється підвищеною стомлюваністю і істощаемостио з нестійкістю настрою, ослабленням самовладання, нетерплячість, непосидючістю, порушенням сну, втратою здатності до тривалої розумової і фізичної напруги, непереносимістю гучних звуків, яскравого світла, різких запахів. Астазія проявляється в здатності м'язів виробляти коливальні і тремтливий руху. М'язовий тремор найбільш виражений на початку і кінці руху, що перешкоджає завершенню цілеспрямованого руху. Видалення мозочка впливає на виконання довільних рухів. Частіше страждають рухи співдружності - симптоми асинергии, в результаті чого відбувається розпад рухів.
У мозочкових хворих деформується хода, т. Е. Виявляється атаксія. Атаксична хода характеризується широко розставленими ногами і надмірними рухами. Порушення координації швидких цілеспрямованих рухів при видаленні або травмах мозочка може викликати адіадокіпез - нездатність виконувати швидку послідовність рухів, наприклад згинання та розгинання пальців. У перенесли травму мозочка через якийсь час настає ефективна компенсація його функцій, здійснювана за рахунок функції кори великих півкуль, яка володіє двосторонніми зв'язками з мозочком. Це свідчить про високу пластичності мозкових систем регулювання руху.
Середній мозок - відділ головного мозку, древній зоровий центр, лежить кпереди від варолиева моста.
У складі середнього мозку розрізняють дві ніжки і четверохолмие. Порожнина середнього мозку представляє вузьку щілину і називається водопроводом мозку. біла речовина ніжок мозку складається з нервових волокон, висхідних (чутливих) і низхідних (рухових) провідних шляхів. Сіра речовина знаходиться всередині і представлено ядрами, найбільшими з яких є чорна речовина і червоне ядро. Червоне ядро парне. У ніжках мозку лежать ядра III і IV пар черепно-мозкових нервів. Від червоних ядер починається спадний проводить шлях, що з'єднує їх з передніми рогами СМ (руброспінальний шлях). Четверохолміе має чотири піднесення-горбка: два верхніх і два нижніх. У них закладені скупчення нервових клітин - ядра. До верхніх ядер четверохолмія підходить частина волокон зорового шляху, до нижніх - слухового. Від ядер четверохолмія йдуть нервові волокна до передніх рогів СМ.
функції середнього мозку різноманітні. Ядра четверохолмия є центрами орієнтовних рефлексів, регулюючи складні рухи тіла при раптових світлових і звукових подразненнях. У середньому мозку відбувається замикання дуги зрачкового рефлексу (звуження зіниці при яскравому освітленні). Ядра ніжок мозку беруть участь в регуляції тонусу м'язів і його розподіл, регулюють ступінь напруги різних труп м'язів, наприклад згиначів і розгиначів.
Проміжний мозок розташовується між середнім і кінцевим мозком і складається з двох частин - таламуса і гіпоталамуса.
таламус (Зорові горби) - парне утворення, що займає основну частину проміжного мозку. Складається з 120 пар ядер, які утворюють три зони: передню; латеральну; медіальну. Таламус має двосторонні зв'язки зі СМ, ретикулярною формацією стовбура мозку, гіпоталамусом, підкірковими ядрами і корою головного мозку. Відповідно до функцій розрізняють специфічні і неспецифічні ядра таламуса. специфічні ядра являють собою переключательние (сенсорні і несенсорні) і асоціативні ядра. Аксони клітин ядер таламуса підходять до певних ділянок кори. Переключательние ядра отримують афференти від різних сенсорних систем або інших відділів мозку і направляють афференти до проекційним зонам кори. В асоціативних ядрах закінчуються афференти від інших таламичних ядер, аксони йдуть до асоціативне зонам кори. неспецифічні ядра не мають специфічних аферентних зв'язків з сенсорними системами, а їх афференти йдуть дифузно до багатьох ділянках кори. Переключательние ядра зорової та слухової сенсорних систем - ядра латерального і медіального колінчастих тіл, соматосенсорної системи - заднє вентральне ядро таламуса. Асоціативними ядрами є латеральні і медіальні ядра подушки. Неспецифічні ядра зосереджені переважно в латеральної, медіальної і середній групах ядер таламуса. Таламус пов'язаний з усіма відділами ЦНС.
Таламус бере участь в переробці сенсорних стимулів, що йдуть до кори великих півкуль, регулює цикл неспання - сон. Вважається, що таламус відіграє важливу роль в здійсненні процесів запам'ятовування. Пошкодження таламуса може привести до антероградной амнезії, викликати тремор - мимовільну тремтіння кінцівок в стані спокою, відсутню, коли пацієнт виконує руху усвідомлено. З таламуса пов'язано рідкісне захворювання, зване «фатальна сімейна безсоння».
гіпоталамус - філогенетично старий відділ проміжного мозку, який грає важливу роль в підтримці сталості внутрішнього середовища і забезпечує інтеграцію функцій вегетативної, соматичної і ендокринної систем. У гіпоталамусі виділяють кілька десятків ядер, топографічно підрозділяються на п'ять груп: 1) преоптична (Перівен- трікулярное, медіальне і латеральне преоптичне); 2) передня (Супрахіазматічне, супраоптичне, паравентрикулярное). Відростки нейронів супраоптичного і іаравентрікулярного ядер йдуть до задньої долі гіпофіза - в нейрогипофиз; 3) середня (Вентромедіальної і дорсоме- діальной ядра); 4) зовнішня (Латеральне гипоталамичне ядро, ядро сірого бугра); 5) задня (Заднє гипоталамичне ядро, періфорнікалиюе ядро, маміллярних ядра).
Середня група ядер формує медіальний гіпоталамус, в якому розташовані нейрони-датчики, що реагують на зміну внутрішнього середовища організму: температуру крові, водно-електролітний склад плазми, вміст гормонів в крові. Медіальний гіпоталамус управляє діяльністю гіпофіза за допомогою нервових і гуморальних механізмів. латеральний гіпоталамус являє собою без'ядерну зону, в якій проходять провідні шляхи (медіальний пучок переднього мозку) до верхніх і нижніх відділах стволової частини мозку.
гіпоталамус отримує афферентний вхід від ретикулярної формації середнього мозку; від середнього мозку; має двосторонні зв'язки з таламус;
зв'язку з корою великих півкуль; афферентний вхід від лімбічної системи мозку; від базальних гангліїв переднього мозку; непрямі мозжечково-гіпоталамічні і вагосупраоптічні зв'язку.
Основні еферентні зв'язку гіпоталамуса з вегетативними і соматичними ядрами головного і СМ здійснюються через полісінаітічні шляху формації стовбура; короткі шляхи в таламус; через таламус в кору великих півкуль; лімбічну кору; середній мозок; нейрогипофиз.
У гіпоталамусі виділяють дві функціонально диференційовані зони - ерготропную і трофотропное. В ерготропной зоні локалізовані вищі центри соматичної нервової системи. Ця зона включає задню і латеральну області гіпоталамуса. Її роздратування викликає соматичні ефекти - розширення зіниць, підвищення артеріального тиску, збільшення частоти серцевих скорочень, припинення перистальтики кишечника; руйнування - тривале зниження тонусу симпатичної нервової системи. при подразненні трофотроп- ної зони спостерігаються ознаки загального збудження парасимпатичної нервової системи, спрямовані на відновлення та збереження резервів організму. Гіпоталамус не є об'єднанням ерготропной і трофот- Ріпне зон. Так, судинозвужувальний ефект може бути отриманий при подразненні і задньої, і передній його областей, т. Е. Симпатичні нейрони присутні в кожній з них.
Функціональна роль гіпоталамуса:
Важливо відмітити
Харчова поведінка являє собою складний комплекс реакцій. Ще в 1912 р видатний американський фізіолог У. Кеннон виявив, що при позбавленні людини або тварини їжі в шлунку починаються ритмічні скорочення, деякі з них можуть супроводжуватися неприємними відчуттями (один з елементів почуття голоду). В цей же час французький патолог Г. Руссі, проводячи дослідження на тваринах, виявив, що при пошкодженні гіпоталамуса підвищується апетит, в деяких випадках спостерігається ненажерливість. У 1951 р американські вчені Дж. Бробек і Б. Ананд знайшли в гіпоталамусі ділянки, при руйнуванні яких тварина припиняло є, - латеральні центри. Надалі американський дослідник Ж. Майер (1952) виявив ділянки головного мозку, які контролюють голодні скорочення шлунка, - центри насичення. У процесі вивчення даного питання виявлено, що почуття насичення з'являється відразу ж після прийому їжі. Надалі Ж. Майер показав, що деякі ділянки головного мозку, названі центрами насичення, контролюють голодні скорочення шлунка і гальмують активність латеральних центрів.
Кінцевий мозок - верхній відділ головного мозку, що досягає самого складної будови у приматів, включаючи людини. Складається з освічених білою речовиною внутрішніх ядер (смугасте тіло і мигдалеподібної комплекс) і поверхнево розташованого сірої речовини - кори великих півкуль, або кори великого мозку.
Площа кори великих півкуль головного мозку людини становить близько 1600 см2, товщина в різних відділах - від 1,3 мм до 4,5 мм. Обсяг кори великих півкуль у людини становить в середньому 44% від обсягу всього півкулі в цілому. Площа поверхні кори однієї півкулі в дорослої людини в середньому дорівнює 220 тис. Мм2. На поверхневі частини припадає 1/3, на що залягають в глибині між звивинами - 2/3 всієї площі кори.
Кора великих півкуль - філогенетично найбільш молодий відділ мозку, в якому відбуваються обробка сенсорної інформації, формування рухових команд і інтеграція складних форм поведінки. Покриває поверхню півкуль і утворює велику кількість різних але глибині і протяжності борозен. Між борознами розташовані різної величини звивини великого мозку. Виділяють п'ять областей (рис. 2.10) - лобову, тім'яну, потиличну, скроневу, примикають
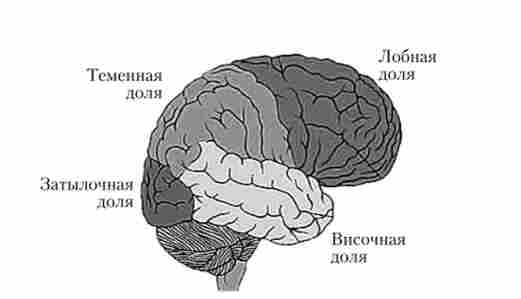
Мал. 2.10. Області кори головного мозку
до відповідних кісток склепіння черепа, і островковую (Острівець), розташовану в глибині латеральної ямки великого мозку, що відокремлює лобову частку від скроневої.
Клітини кори за морфологічним ознакою поділяються на кілька груп. пірамідні клітини орієнтовані вертикально по відношенню до поверхні кори і мають тіло трикутної форми. Від тіла відходить вгору довгий Т-образно розгалужених апікальний дендрит, вниз - аксон, який або йде з кори в складі низхідних шляхів, або до інших зон кори. Апікальні і більш короткі базальні дендрити пірамідних клітин усіяні дрібними виростами - шипиками, які представляють собою область синаптичного контакту. зірчасті клітини - більш дрібні, мають короткі розгалужені дендрити і аксони, що формують внутрікорті- Кальне зв'язку. веретеновідние клітини мають довгий аксон, орієнтований в горизонтальному або вертикальному напрямку.
Кора ссавців побудована по екранному принципу (тіла і відростки мають впорядковане будова) і в типовому випадку складається з шести горизонтальних шарів.
На підставі відмінностей в будові па клітинному рівні (цитоархітектоніка) відділів кори великих півкуль її прийнято розділяти на поля (цітоархітектонічно поля Бродмана). У 1909 році німецьким неврологом К. Бродманом на зображенні (карті) зовнішньої і внутрішньої поверхні півкуль головного мозку були відзначені 52 поля.