| Головна |
| «« | ЗМІСТ | »» |
|---|
Нервова регуляція кровообігу вирішує кілька найважливіших для існування організму завдань. По-перше, вона підтримує такий рівень артеріального тиску, при якому забезпечується повноцінне кровопостачання ГМ і серця. По-друге, за умови виконання першого завдання, постачає кров'ю всі інші органи. По-третє, за умови виконання двох перших завдань, здійснює перерозподіл органного кровотоку у відповідності з поточними фізіологічними завданнями організму (доповнюючи місцеві і гуморальні механізми). Перераховані завдання вирішуються за допомогою ВНС, здатної ефективно впливати на насосну функцію серця (див. Гл. 6) і на тонус судин всіх органів і тканин.
Нервова регуляція судин здійснюється ВНС, головним чином - її симпатичним відділом. Вегетативні нерви іннервують всі судини, крім капілярів (у них немає гладких м'язів - основний мішені нервової регуляції).
Щільність нервових закінчень в різних частинах судинної системи неоднакова: артерії, як правило, іннервіровани краще, ніж вени, а артеріоли і дрібні артерії - краще, ніж великі. Нервові закінчення лежать на кордоні гладеньком'язового шару і зовнішньої оболонки, де вони розгалужуються, утворюючи синаптичні здуття (варикоз). Розташування нервових закінчень в стінці артеріоли і схема іннервації нреканіллярних судин показані на рис. 8.5.
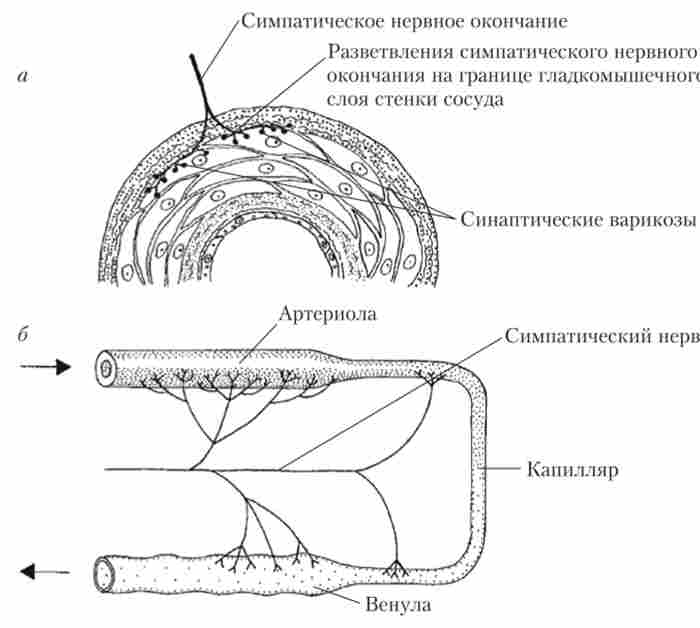
Мал. 8.5. Схема розташування нервових закінчень в стінці судини (а); іннервація мікросудин (Б)
Симпатичні нервові закінчення в більшості випадків виділяють в синаптичну щілину НА, що викликає скорочення гладкої мускулатури стінок судин. За симпатичним нервовим закінченням до судин безперервно йде потік імпульсів з частотою 1-3 в секунду, який викликає скорочення їх гладких м'язів. Це скорочення підтримує судини в частково судженого стані (нейрогенний судинний тонус). Зменшення імпульсації по симпатичних судинозвужувальних нервів призводить до розширення судин, посилення іміульсаціі - до звуження судин. Таким чином, наявність нейрогенного тонусу розширює регуляторні можливості симпатичної нервової системи.
У деяких видів тварин, наприклад у собак і кішок, існує особлива система симпатичних волокон, иннервирующая тільки артеріальні судини скелетних м'язів. Ця система активується при стимуляції гіпоталамуса. У спокої імпульсація в цих волокнах відсутня, але з'являється при емоційних реакціях: люті, страху, тривозі і ін. Активація цих волокон призводить до викиду з нервових закінчень АХ, що викликає розширення судин. Значення цієї реакції полягає в підготовці м'язів до майбутньої роботи. У людини такі волокна поки не виявлені, хоча розширення судин скелетних м'язів, що передує фізичному навантаженні, у нього спостерігається.
Судини зовнішніх статевих органів іннервуються волокнами парасимпатичного відділу ВНС. У спокої імпульсація в цих волокнах відсутня, але з'являється при статевому збудженні. Закінчення цих волокон виділяють АХ і NO, який розширює судини і збільшує кровонаповнення статевих органів. Парасимпатичну іннервацію мають також судини легких і ГМ.
Таким чином, провідна роль в регуляції тонусу судин належить симпатичному відділу ВНС. Роль парасимпатичного відділу незначна. Це відрізняє нервову регуляцію судин від регуляції серця, де обидва відділи нервової системи однаково важливі.
Прегангліонарних симпатичні нейрони, які беруть участь в нервової регуляції серцево-судинної системи, лежать в бічних рогах сірої речовини великої групи сегментів спинного мозку (від восьмого шийного до третього поперекового) і посилають свої аксони до ганглиям, що створює симпатичні ланцюжка (стовбури). У ланцюжках ці аксони можуть утворювати синапси на нейронах, що лежать в різних гангліях. Симпатичні гангліонарні нейрони безпосередньо іннервують серце і судини, причому в стінці однієї судини зазвичай розташовуються нервові закінчення, пов'язані з нейронами з декількох гангліїв. Основні шляхи ефек- торного ланки нервової регуляції серцево-судинної системи показані на рис. 8.6.
Активність прегангліонаріих симпатичних нейронів перебуває під контролем нейронної мережі, розташованої в довгастому мозку і вароліевом мосту. Ця мережа отримала назву серцево-судинного центру.
Серцево-судинний центр ГМ розташовується в ретикулярної формації довгастого мозку і нижньої третини моста. У його склад входить ядро блукаючого нерва і зони, активація яких викликає звуження судин з одночасним посиленням роботи серця (прессорний центр) і розширення судин з одночасним пригніченням роботи серця (депрес- бур'янистої центр). Нейрони прессорного центру активують симпатичні прегангліонарних нейрони і гальмують парасимпатичні нейрони ядра блукаючого нерва. Нейрони депрессорного центру гальмують активність нейронів прессорного центру і активують нейрони ядра блукаючого нерва.
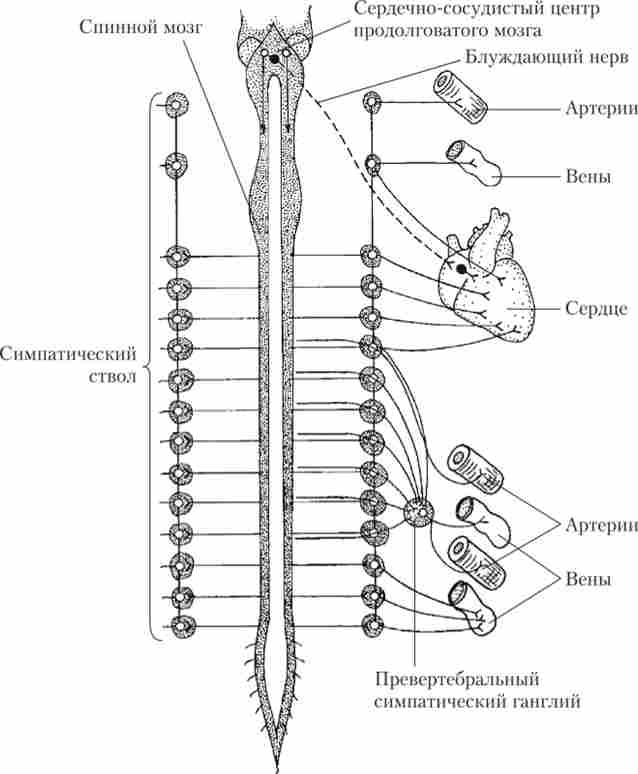
Мал. 8.6. Схема анатомічної будови основних шляхів і центрів симпатичної регуляції серцево-судинної системи:
пунктиром показаний парасимпатичний шлях в складі блукаючого нерва
Серцево-судинний центр, з одного боку, отримує і інтегрує інформацію від численних рецепторів серцево-судинної системи, а з іншого - знаходиться під постійним контролем верхніх нервових центрів (рис. 8.7).
На серцево-судинний центр впливають рецептори, що вимірюють системний тиск крові (барорецептори); рецептори, що вимірюють рівень кровопостачання тканин (хеморецептори 02, С02 та ін.); рецептори, що вимірюють стан стінок серця (механорецептори серця), і ін. Серцево-судинний центр взаємодіє з дихальним центром, який також розташовується в структурах довгастого мозку і моста.
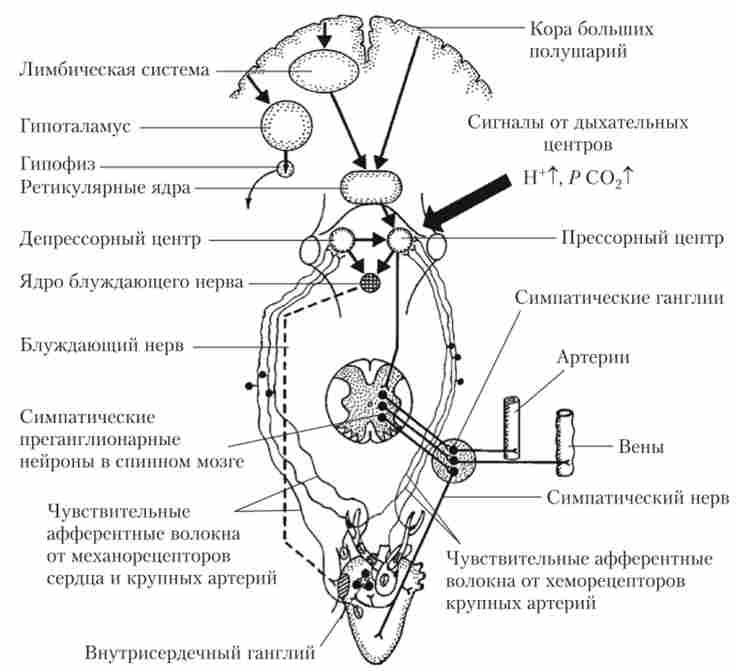
Мал. 8.7. Схема основних зв'язків серцево-судинного регуляторного центру довгастого мозку
З верхніх відділів ГМ на серцево-судинний центр найбільший вплив надають ядра ретикулярної формації моста, середнього мозку і гіпоталамуса. На серцево-судинний центр впливають також лімбічна система і деякі відділи кори великих півкуль: рухова область, передня частина скроневої частки, орбітальна область лобової кори, передня частина поясної звивини і ін. Області ГМ, що впливають на серцево-судинний центр, показані на рис. 8.8.
Більшість з перерахованих структур можуть надавати на роботу серцево-судинного центру як гнітюче, так і стимулюючий вплив в залежності від інтенсивності збудження і вихідної активності центру.
барорецептори, здійснюють контроль за рівнем тиску, розташовуються в основному у великих судинах і в серці. Там вони утворюють скупчення, які називаються рефлексогенних зонами. Найважливішими з них є зони дуги аорти і каротидного синуса (місце розгалуження сонної артерії на внутрішню і зовнішню). Розташування цих зон показано на рис. 8.9.
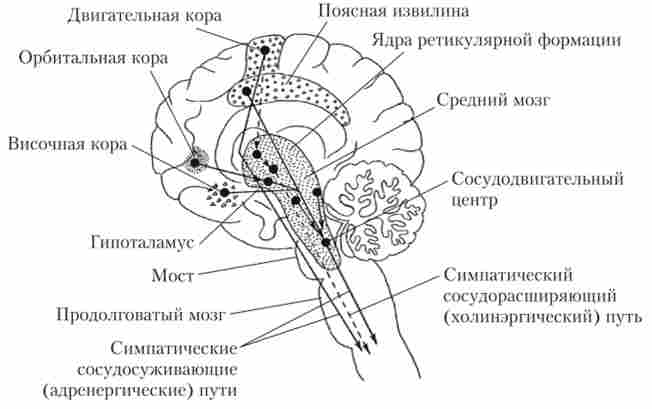
Мал. 8.8. Області головного мозку, що беруть участь в регуляції кровообігу, і зв'язуючі їх шляху
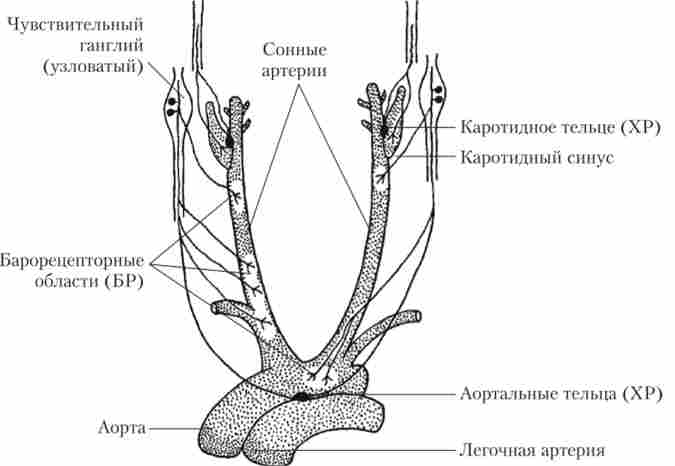
Мал. 8.9. Розташування барорецепторів (БР) і хеморецепторів (ХР) в аорті і сонної артерії
Барорецептори по суті є рецепторами розтягування, т. Е. Механорецепторами, що реагують на зміни напруги в стінці судини. Барорецептори активні вже при нормальному рівні тиску крові. снидружин не тиску під час діастоли шлуночків зменшує їх активність, а підйом тиску під час систоли шлуночків - збільшує. При зростанні тиску вище норми частота імпульсації від барорецепторів прогресивно зростає. Сигнали від барорецепторів надходять в депрес- бур'янистої центр довгастого мозку, який у відповідь гальмує активність прессорного центру і активує блукаючий нерв. В результаті судини розширюються, а серце гальмує свою робіт} '. У сукупності це призводить до дуже швидкого й ефективного зниження тиску. Активність барорецепторів при цьому теж знижується. Навпаки, при зниженні артеріального тиску частота імпульсів, що йдуть від барорецепторів, зменшується, що призводить до рефлекторного посилення роботи серця і звуження судин. В результаті артеріальний тиск підвищується. Таким чином, барорефлексов стабілізують тиск на певному рівні. Цей рівень задається серцево-судинним центром під впливом сигналів від інших рецепторів або верхніх відділів ГМ.
Серцеві механорецептори були описані в гл. 6.
Артеріальні хеморецептори, розташовані в каротидних і аортальних тільцях (див. рис. 8.9), не тільки грають важливу роль в рефлекторної регуляції дихання, але і впливають на систему кровообігу.
Хеморецептори представляють собою клітини, чутливі до нестачі кисню, а також надлишку вуглекислого газу і іонів водню. Ці клітини розташовуються в невеликих (діаметром близько 2 мм) хеморе- цептівних органах. Два з них лежать в області каротидного синуса (каротидні тільця), а два-три - поблизу аорти (аортальні тільця). Імпульси від цих хеморецепторів надходять в серцево-судинний центр по чутливих нервових волокнах, що йде паралельно з волокнами від барорецепторів (див. Рис. 8.9). Хеморецептори знаходяться в постійному контакті з кров'ю, так як густо обплетені капілярами. Коли артеріальний тиск падає, хеморецептори починають відчувати брак кисню і надлишок вуглекислого газу і іонів водню, в результаті чого вони порушуються. Посилена пульсація від рецепторів активує серцево-судинний центр, який повертає тиск до початкового рівня.
Таким чином, комплекс баро-і хеморецепторів здійснює постійний контроль за рівнем артеріального тиску, посилаючи сигнали про його відхилення від норми в серцево-судинний центр. Цей центр, впливаючи на роботу серця і тонус судин, забезпечує рефлекторну стабілізацію тиску. Нейрогенні механізми при необхідності доповнюються гуморальними, зокрема - ендокринними, які також здатні брати участь в регуляції роботи серця і тонусу судини. Час спрацювання нейрогенних механізмів регуляції - секунди, гуморальних - хвилини і годинник, що дозволяє віднести їх до системи швидкої регуляції кровообігу. В організмі, однак, існує потужний механізм регуляції артеріального тиску, ефективність якого проявляється не так швидко, але підтримується протягом тижнів і навіть місяців. Цей механізм може бути віднесений до системи довготривалої регуляції кровообігу.