| Головна |
| «« | ЗМІСТ | »» |
|---|
Викопні елементи клітин еукаріотичного типу виявлені в породах, вік яких не перевищує 1,0-1,4 млрд років. Пізніше виникнення, а також схожість в загальних рисах їх основних біохімічних процесів (самоудвоение ДНК, синтез білка на рибосомах) змушують думати про те, що еукаріотичні клітини походять від предків, які мали прокариотичний будова.
Найбільш популярна в даний час симбиотична гіпотеза походження еукаріотичних клітин, згідно з якою (рис. 1.5, /) основою, або клітиною-господарем, в еволюції клітини еукаріотичного типу послужив анаеробний прокаріот, здатний лише до амебоідному руху. Перехід до аеробного дихання пов'язаний з наявністю в клітині мітохондрій, які відбулися шляхом змін симбіонтів - аеробних бактерій, що проникнули в клітку-господаря і співіснували з нею.
Подібне походження припускають для джгутиків, предками яких служили симбіонти-бактерії, що мали джгутик і нагадували сучасних спирохет. Придбання кліткою джгутиків мало поряд з освоєнням активного способу руху важливий наслідок загального порядку. Припускають, що базальні тільця, якими забезпечені джгутики, могли еволюціонувати в центриоли в процесі виникнення механізму мітозу.
Здатність зелених рослин до фотосинтезу обумовлена присутністю в їх клітинах хлоропластів. Прихильники симбиотичної гіпотези вважають, що симбионтами клітини-господаря, дали початок хлоропластам, послужили прокариотичні синьо-зелені водорості.
Серйозним аргументом на користь симбіотичного походження мітохондрій, центриолей і хлоропластів є те, що перераховані органели мають власну ДНК. Разом з тим білки баціллін і тубулін, з яких складаються джгутики і війки відповідно сучасних прокаріот і еукаріот, мають різну будову. У сучасних бактерій, не знайдено також структур з властивою жгутикам, віям, базальним тільцям і центрі- олям еукаріотів комбінацією микротрубочек: «9 + 2» або «9 + 0».
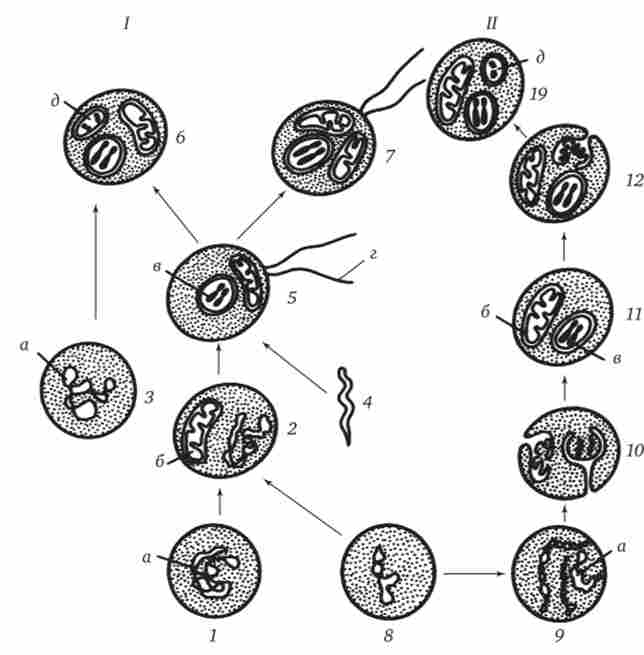
Мал. 1.5. Походження еукаріотичної клітини згідно симбіотичного (Г) і інвагінаціонний (Я) гіпотезам.
I - анаеробний прокаріотів (клітина-господар), 2 - прокаріоти, мають мітохондрії, 3 - синьо-зелених водоростей (презумптівного хлоропласт), 4 - спірохетообразная бактерія (презумптівного джгутик), 5 - примітивний еукаріот зі джгутиком, 6 - рослинна клітина, 7-тваринна клітина зі джгутиком, 8 - аеробний прокаріотів (презумп- тивная мітохондрія), 9 - аеробний прокаріотів (клітина-родоначальниця відповідно до гіпотези Я), 10 - инвагинации клітинної оболонки, що дали ядро і мітохондрії,
II - примітивний еукаріот, 12 - впячивание клітинної оболонки, що дало хлоропласт, 13 - рослинна клітина; а - ДНК клітини прокаріотів, б - мітохондрія, в - ядро еукаріотичної клітини, г - джгутик, д - хлоропласт
Внутрішньоклітинні мембрани гладкою і шорсткою цитоплазматичної сітки, пластинчастого комплексу, бульбашок і вакуолей розглядають як похідні зовнішньої ядерної оболонки, яка здатна утворювати випинання.
Центральним і складним для відповіді є питання про походження ядра. Припускають, що воно також могло утворитися з сімбі- Онта-прокаріоти. Збільшення кількості ядерної ДНК, у багато разів перевищує в сучасній еукаріотичної клітці її кількість в мітохондрії або хлоропласті, відбувалося, мабуть, поступово шляхом переміщення груп генів з геномів симбіонтів. Не можна виключити, однак, що ядерний геном формувався шляхом нарощування генома клітини-господаря (байдужості симбіонтів).
згідно інвагінаціонний гіпотезі, предковой формою еукаріотичної клітини був аеробний прокаріотів (Див. Рис. 1.5, IT). Усередині такої клітини-господаря перебувало одночасно кілька геномів, але спочатку прикріплюється до клітинної оболонці. Органели, що мають ДНК, а також ядро, виникли шляхом впячивания і отшнуро- виванія ділянок оболонки з подальшою функціональною спеціалізацією в ядро, мітохондрії, хлоропласти. У процесі подальшої еволюції відбулося ускладнення ядерного генома, з'явилася система цитоплазматичних мембран.
Інвагінаціонний гіпотеза добре пояснює наявність в оболонках ядра, мітохондрій, хлоропластів, двох мембран. Однак вона не може відповісти на питання, чому біосинтез білка в хлоропла- стах і мітохондріях в деталях відповідає такому в сучасних прокариотичних клітинах, але відрізняється від біосинтезу білка в цитоплазмі еукаріотичної клітини.
еволюційні можливості клітин еукаріотичного типу незрівнянно вище, ніж прокариотичного. Провідна роль тут належить ядерного геному еукаріот, який у багато разів перевершує за розмірами геном прокаріотів. Кількість генів у бактерії і в клітці людини, наприклад, співвідноситься як 1: (100 + 1000). Важливі відмінності полягають в диплоидности еукаріотів завдяки наявності в ядрах двох комплектів генів, а також в багаторазовому повторенні деяких генів. Це розширює масштаби мутаційної мінливості без загрози різкого зниження життєздатності, еволюційно значущим наслідком чого є утворення резерву спадкової мінливості.
При переході до еукаріотичних типу ускладнюється механізм регуляції життєдіяльності клітини, що на рівні генетичного матеріалу проявилося в збільшенні відносної кількості регуляторних генів, заміні кільцевих «голих» молекул ДНК прокаріот хромосомами, в яких ДНК з'єднана з білками. В результаті стало можливим зчитувати біологічну інформацію по частинах з різних груп генів в різному їх поєднанні в різних типах клітин і в різний час. У бактеріальній клітині, навпаки, одночасно зчитується до 80-100% інформації генома. У клітинах дорослої людини в різних його органах транскрибується від 8-10% (печінка, нирка) до 44% (головний мозок) інформації. Використання біологічної інформації частинами належить виняткова роль в еволюції багатоклітинних організмів, так як саме це дозволяє різним групам клітин спеціалізуватися за різними функціональними напрямками.
Велике значення при переході до многоклеточности мало наявність у еукаріотів еластичною оболонки, що необхідно для утворення стійких клітинних комплексів.
Серед цитофізіологічної особливостей еукаріот, що збільшують їх еволюційні можливості, необхідно назвати аеробне дихання, яке також стало причиною для розвитку багатоклітинних форм. еукаріотичні клітини з'явилися на Землі після того, як концентрація 02 в атмосфері досягла 1% (точка Пастера). Названа концентрація є необхідною умовою аеробного дихання.
В умовах ускладнення генетичного апарату еукаріот, збільшення сумарної кількості ДНК і розподілу її по хромосомах важко переоцінити значення виникнення в еволюції митоза як механізму відтворення в поколіннях генетично подібних клітин.
Поява внаслідок еволюційних перетворень мітозу такого способу ділення клітин, як мейоз, що дає можливість зберегти сталість хромосом в ряду поколінь, найкращим чином вирішило проблему розмноження багатоклітинних організмів. Пов'язаний з мейозом перехід до статевого розмноження посилив еволюційну роль комбинативной мінливості, сприяв збільшенню швидкості еволюції.
Завдяки зазначеним особливостям за 1 млрд років еволюції еукаріотичний тип клітинної організації дав широке розмаїття живих форм від одноклітинних найпростіших до ссавців і людини.