| Головна |
| «« | ЗМІСТ | »» |
|---|
У зв'язку з особливостями організації окремих генів еукаріот і геному в цілому регуляція генної активності у них характеризується деякими відмінностями в порівнянні з прокариотами.
У еукаріот не встановлено оперон організації генів. Гени, що визначають синтез ферментів одного ланцюга біохімічних реакцій, можуть бути розсіяні в геномі і, очевидно, не мають, як у прокаріотів, єдиної регулюючої системи (ген-регулятор, оператор, промотор). У зв'язку з цим синтезовані мРНК у еукаріот моноцістронни, т. Е. Є матрицями для окремих пептидних ланцюгів.
В даний час механізми регуляції і координації активності еукаріотичних генів інтенсивно вивчаються. Встановлено, що їх функціонування безсумнівно підпорядковується регуляторним впливам, проте регуляція транскрипції в еукаріот є комбінаційної, т. е. активність кожного гена регулюється великим спектром генів-регуляторів (рис. 3.87).
У багатьох еукаріотичних генів, що кодують білки і транскрібіруемих РНК-полімераза II, в ДНК є кілька областей, які впізнаються різними білками-регуляторами. Однією з них є область, розташована поблизу промотора. Вона включає близько 100 пар нуклеотидів, в тому числі ТАТА-блок, розташований на відстані 25 пар нуклеотидів від точки початку транскрипції. Встановлено, що для успішного приєднання РНК-полімерази II до промотор необхідно попереднє з'єднання з ТАТА-блоком особливого білка - фактора транскрипції - з утворенням стабільного транскрипційного комплексу. Саме цей комплекс ДНК з білком впізнається РНК-полімераза II. Послідовності нуклеотидів, що примикають до ТАТА-блоку, формують необхідний для транскрипції елемент, який закриває передню частину промотором.
Інша область, яка відіграє важливу роль в регуляції активності еукаріотичних генів, розташовується на великій відстані від промотора (до декількох тисяч пар нуклеотидів) і називається енхансером (Від англ, enhance - посилювати).
І енхансер, і препромоторний елемент еукаріотичних генів містять серію коротких нуклеотидних послідовностей,
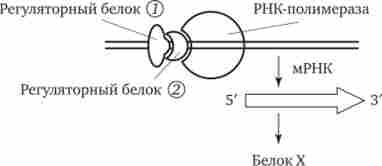
Мал. 3.87. Регуляція експресії гена, що кодує білок X у еукаріот, двома регуляторними білками які зв'язуються з відповідними регуляторними білками. В результаті взаємодії цих білків відбувається включення або виключення генів.
Особливістю регуляції експресії еукаріотичних генів є також існування білків-регуляторів, які здатні контролювати транскрипцію багатьох генів, що кодують, можливо, інші білки-регулятори. У зв'язку з цим деякі (головні) білки-регулятори мають координуючим впливом на активність багатьох генів і їх дія характеризується плей отропним ефектом (рис. 3.88). Прикладом може служити існування білка, який активує транскрипцію декількох специфічних генів, що визначають диференціювання попередників жирових клітин.
З огляду на те що в геномі еукаріотів є багато надлишкової ДНК, а в кожній клітині організму транскрибується всього 7-10% генів, логічно припущення про те, що у них переважає позитивний генетичний контроль, при якому активація невеликої частини генома виявляється більш економічною, ніж репресія основний маси генів.
Безперечною особливістю регуляції транскрипції в еукаріот є підпорядкованість цих процесів регулюючим впливам з боку гормонів організму. Останні часто грають роль індукторів транскрипції. Так, деякі стероїдні гормони оборотно зв'язуються особливими білками-рецепторами, утворюючи з ними комплекси. Активоване гормоном рецептор набуває здатності з'єднуватися зі специфічними ділянками хроматину, відповідальними за регуляцію активності генів, в яких рецептори дізнаються певні послідовності ДНК.
Специфічність регулюючого впливу гормону на транскрипцію обумовлена не тільки природою самого гормону, а й природою клітини-мішені, що синтезує специфічний білок-рецептор, який впливає на транскрипцію певного для даної клітини набору генів. Прикладом участі гормонів в регуляції активності певних генів може служити вплив тестостерону на розвиток тканин організму за чоловічим типом при наявності специфічного білка-рецептора. Відсутність останнього при мутації відповідного гена не дає можливості гормону проникнути в ядра клітин-мішеней і забезпечити включення певного набору генів: розвивається синдром тестикулярной фемінізації, або синдром Морріса (див. Розд. 3.6.5.2).
Наступна особливість регуляції генної активності у еукаріот пов'язана з утворенням стійкого комплексу ДНК з белкамі-хроматину (Див. Розд. 3.5.2.2). Провідна роль в компактизации ДНК належить гістонів, тому вони, безсумнівно, беруть участь і в процесах регуляції генної активності (див. Розд. 3.5.4). Неодмінною умовою для здійснення транскрипції в еукаріот є попередня декомпактізаціі хроматину на відповідній ділянці,
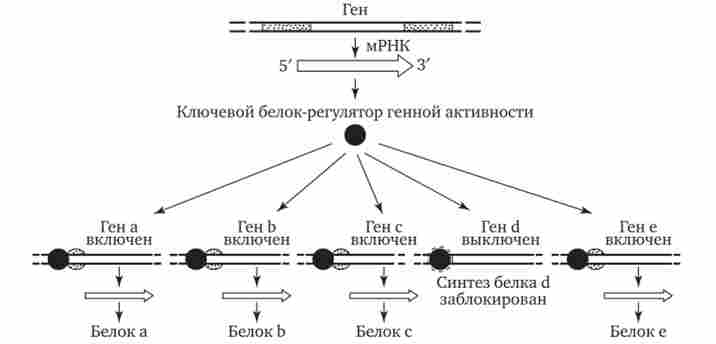
Мал. 3.88. Регуляція експресії багатьох генів еукаріот одним білком-регулятором
де тимчасово втрачається зв'язок з Н, -гістонамі і кілька послаблюється зв'язок з нуклеосомної гистонами. Правда, нуклеосомна організація хроматину не втрачається навіть в ході транскрипції, проте контакт ДНК і негістонових білків стає можливим і відбувається дерепресія гена.
Відмінною особливістю регуляції експресії генів у еукаріот є можливість її здійснення не тільки на стадії транскрипції, а й на інших етапах розтягнутого в часі процесу реалізації спадкової інформації. Регуляція на стадії транскрипції є найбільш економічною, але недостатньо швидко реагує на зміну ситуації. Так, виникла в клітці потреба в будь-якому білку не може бути швидко задоволена шляхом включення транскрипції відповідного гена. Синтезований транскрипт повинен піддатися процесингу, потім зріла мРНК повинна вийти з ядра в цитоплазму і, утворюючи комплекс з рибосомами, здійснити трансляцію інформації, синтезувавши пептид, який, лише пройшовши посттрансляційних зміна, формує активний білок, необхідний клітці.
У тому випадку, коли клітці потрібно припинити синтез якогось продукту, після виключення транскрипції відповідного гена в цитоплазму деякий час будуть продовжувати надходити дозрівають молекули мРНК, які здійснюють там синтез пептидних ланцюгів, поки вони не деградують під дією ферментів. Таким чином, для ефективного регулювання експресії генів у еукаріот повинні існувати механізми, що працюють не тільки на стадії транскрипції, а й на інших етапах цього процесу.
Пов'язана з екзон-інтронів організацією генів необхідність процесингу, в тому числі сплайсингу, уможливлює регуляцію цих процесів в ядрі. В даний час обговорюється роль інтронних ділянок ДНК в зміні схеми сплайсингу при синтезі антитіл (див. Розд. 3.4.3.2) або цитохрому b (Див. Розд. 3.4.3.3). Це створює можливість, використовуючи один і той же первинний транскрипт, забезпечувати освіту матриць для різних пептидів, вирізаючи з них різні послідовності або змінюючи послідовності на S'- і З'-кінцях мРНК.
Очевидно, і транспорт зрілих мРНК з ядра в цитоплазму також регулюється певним чином, так як встановлено, що лише невелика частина РНК, транскрибируемой з генів, після сплайсингу залишає ядро. Значна кількість її деградує. Можливо, це є результатом процесингу, що спричиняє появу «неправильних» матриць.
Існують механізми, що забезпечують регуляцію процесів синтезу пептидних ланцюгів. Вони менш економічні, але відрізняються швидкістю реагування на зміни потреб клітини в даному білку. Регуляція трансляції здійснюється на стадії ініціації шляхом впливу на один з факторів ініціації, що каталізує приєднання до малої субодиниці рибосоми тРНК, що несе метіонін (формілметіонін) (див. Розд. 3.4.3). В результаті при наявності в цитоплазмі мРНК трансляції на ній не відбувається. Така ситуація спостерігається, наприклад, при відсутності в цитоплазмі гема, що веде до виключення трансляції глобінових ланцюгів гемоглобіну.
Нарешті, регуляція процесу реалізації спадкової інформації може здійснюватися і на стадії посттрансляційних змін. Припинення цих процесів обумовлює затримку у формуванні активних молекул білка при наявності необхідних для цього пептидних ланцюгів. Наприклад, для формування активної форми білкового гормону - інсуліну - з проінсуліну повинні вирізатися дві субодиниці. Гальмування цих процесів зменшує вихід кінцевого активного продукту.
Таким чином, розглянутий вище приклад регуляції експресії генів демонструє складні взаємозв'язки, які існують між ними в геномі. Формування будь-якої ознаки тому не можна розглядати як результат дії однієї пари алельних генів в генотипі. У будь-якому випадку регуляція експресії відповідального за цю ознаку гена здійснюється за участю інших генів.