Вивчення регуляції генної активності у прокаріотів призвело французьких мікробіологів Ф. Жакоба і Ж. Моно до створення (1961) оперон моделі регуляції транскрипції. оперон - це тісно пов'язана послідовність структурних генів, що визначають синтез групи білків, які беруть участь в одного ланцюга біохімічних перетворень. Наприклад, це можуть бути гени, які детермінують синтез ферментів, які беруть участь у метаболізмі певної речовини або в синтезі якогось компонента клітини. Оперон модель регуляції експресії генів передбачає наявність єдиної системи регуляції у таких об'єднаних в один оперон структурних генів, що мають загальний промотор і оператор.
Особливістю прокариот є транскрибування мРНК з усіх структурних генів оперона у вигляді одного поліцістронной транскрипта, з якого в подальшому синтезуються окремі пептиди.
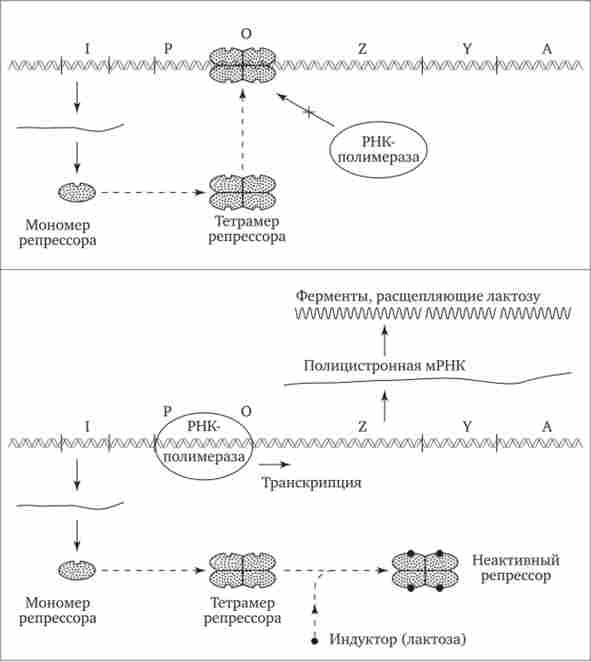
Прикладом участі генетичних та негенетичних чинників в регуляції експресії генів у прокаріотів може служити функціонування лактозного оперона у кишкової палички Е. coli (Рис. 3.86). При відсутності в середовищі, на якій вирощуються бактерії, цукру лактози активний білок-репрессор, який синтезується геном-регулятором (I), взаємодіє з оператором (О), перешкоджаючи з'єднанню РНК-полімерази з промотором (Р) і транскрипції структурних
Мал. 3.86. лактозна оперон Е. coli.
Включення (ас-оперону за допомогою індуктора - лактози; пояснення см. В тексті
генів Z, Y, А. Поява в середовищі лактози інактивує репрессор, він не з'єднується з оператором, РНК-полімераза взаємодіє з промотором і здійснює транскрипцію поліцістронной мРНК. Остання забезпечує синтез відразу всіх ферментів, які беруть участь у метаболізмі лактози. Зменшення вмісту лактози в результаті її ферментативного розщеплення призводить до відновлення здатності репрессора з'єднуватися з оператором і припинення транскрипції генів Z, Y, А.
Таким чином, регуляція експресії генів, організованих у прокаріотів в Оперон, є координованої. Синтез поліцістронной мРНК забезпечує однаковий рівень синтезу всіх ферментів, які беруть участь в біохімічному процесі.
Ретикулярна формація - нервова система: анатомія, фізіологія, Нейрофармакологія Виходячи зі сказаного в попередньому параграфі, виникає питання: якщо в довгастому мозку і мосту нейрони рогів СМ перетворюються в ядра черепних нервів, то чому ж стає його проміжне ядро? Нагадаємо, що ця область є найбільш медіальної частиною сірої речовини СМ і найважливішим центром, який
Реплікація ДНК - генетика в 2 Ч. Частина 1 Розшифровка структури молекули ДНК допомогла пояснити і принцип її реплікації (подвоєння) в клітці. Цей принцип полягає в тому, що кожна з двох полінуклеотидних ниток молекули ДНК служить в якості програми (матриці) для синтезу нової (комплементарної) нитки. В результаті на основі
Рекомбінаційний механізм хромосомних перебудов - генетика Вперше припущення про рекомбінаційному механізмі хромосомнихаберацій висловив А. С. Серебровський (1929). Він виходив з реципрокного характеру транслокаций, рекомбінаційного походження дуплікацій і делеций, а також передбачав необхідність реципрокного обміну в разі інверсій. Було відмічено
Регуляція циклу трикарбонових кислот - біохімія Основними регулярними ферментами циклу є ферменти, що каталізують практично незворотні реакції: цітратсінтаза (реакція (I)] і ізоцитратдегідрогеназа (реакція (3) (. Відомими інгібіторами першого ферменту є АТФ, НАДН і сукцинил-SKoA. Однак вважають, що головну регуляторну функцію в цьому процесі
Регуляція синтезу білка, регуляція синтезу білка у прокаріотів - біохімія частина 2. Синтез білка - складний, багатоступінчастий процес, який залежить від функціонального стану ДНК, РНК і безпосередньо білок-сінтезіруюшей системи. Тому механізми регуляції швидкості утворення білка реалізуються як в ядрі, так і в цитоплазмі. З розглянутого зрозуміло, що в освіті поліпсптідной
Регуляція глюконеогенезу - біохімія Регуляторним ферментом в глюконеогенезі є піруваткарбоксіла- за, що каталізує першу необоротну реакцію цього процесу. Позитивним аллостеричним ефектором ферменту (активатором) є ацетил-КоА. Тому біосинтез глюкози відбувається тоді, коли в мітохондріях накопичується більше ацетил-КоА, ніж потрібно