| Головна |
| «« | ЗМІСТ | »» |
|---|
Розшифровка структури молекули ДНК допомогла пояснити і принцип її реплікації (подвоєння) в клітці. Цей принцип полягає в тому, що кожна з двох полінуклеотидних ниток молекули ДНК служить в якості програми (матриці) для синтезу нової (комплементарної) нитки. В результаті на основі однієї двухцепочечной молекули утворюються дві однакові дволанцюжкові молекули, в кожній з яких один ланцюжок є старої, а інша - нової (знову синтезованої). Такий принцип реплікації ДНК був названий напівконсервативним (рис. 5.12). Відповідно до цього принципу нуклеотидних послідовність матричної (батьківської) нитки зчитується в напрямку 3 '-> 5', тоді як синтез нової (дочірньої) нитки йде в напрямку 5'? З '. Оскільки дві комплементарні ланцюжка батьківської молекули ДНК є антипаралельними, то синтез нової полинуклеотидной ланцюжка на кожній з них йде в протилежному напрямку.
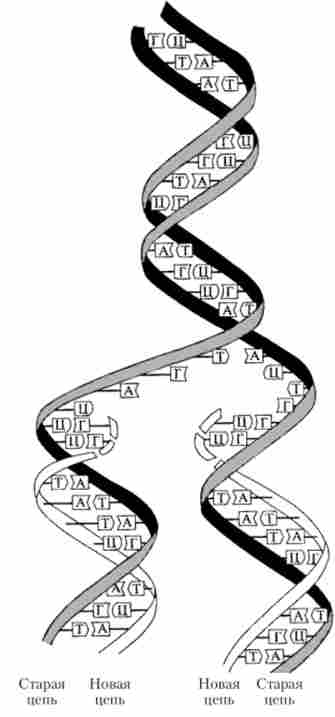
Мал. 5.12. Принцип напівконсервативної реплікації ДНК
Механізм реплікації ДНК є досить складним і, ймовірно, різниться в разі організмів, що містять відносно невеликі за розмірами молекули ДНК в замкнутій (кільцевій) формі (багато вірусів і бактерії), і еукаріот, клітини яких мають молекули величезних розмірів, що знаходяться в лінійній (незамкненою ) формі.
Невелика кільцева молекула ДНК являє собою одну структурну одиницю реплікації (реплікон), що має єдину точку початку (ініціації) реплікації (0-пункт, що складається приблизно з 300 нуклеотидів), в якій починається процес розбіжності (розплітання) двох ниток батьківської молекули і матричного синтезу комплементарних копій (реплік) дочірньої ДНК. Цей процес триває безперервно по довжині копируемой структури і закінчується в цьому ж РЕПЛІКОН утворенням двох молекул «напівконсервативним» типу. У великих лінійних молекулах ДНК еукаріот є багато точок початку реплікації і відповідних їм репліконов (від декількох сотень до десятків тисяч), т. Е. Така ДНК є полірепліконной.
При розгляді сучасних уявлень про механізм реплікації ДНК еукаріот можна умовно виділити гри послідовні етапи цього процесу, що відбувається в РЕПЛІКОН, в кожному з яких беруть участь ті чи інші білки (ферменти).
Перший етап пов'язаний з швидким розкручуванням двох полінуклеотидних ниток спіраль молекули ДНК на певному її ділянці (в межах працюючого реплікону) і з їх поділом шляхом руйнування водневих зв'язків між парами комплементарних основ. При цьому утворюються два одноланцюгових фрагмента батьківської молекули, кожен з яких може виступила пать в ролі матриці для синтезу комплементарної (дочірньої) нитки. Цей етап ініціюється у відповідній точці розпочато реплікації і забезпечується комплексним участю декількох різних білків. В результаті їх дії формується У-образна структура, названа вилкою реплікації, в якій дві батьківські ланцюжка ДНК вже відокремлені один від одного (рис. 5.13). Новоутворена вилка реплікації швидко просувається уздовж подвійної спіралі батьківської молекули ДНК завдяки активності «розплітає» ферменту ДНК-гслікази і за участю групи дестабілізуючих білків. Ці білки мають здатність зв'язуватися тільки з одноланцюжковий (вже розкрученими і розділеними) ділянками молекули, перешкоджаючи виникненню на них вторинних складчастих утворень ( «шпильок») за рахунок випадкових з'єднань між комплементарними нуклеотидами одноні- тєвої структури. Отже, вони сприяють випрямляння одноні- тевьгх ділянок молекули, що необхідно для нормального виконання ними матричних функцій.
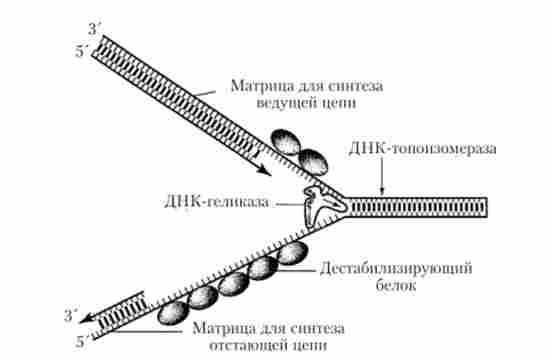
Мал. 5.13. Схема освіти вилки реплікації ДНК
Швидке розплітання ДНК за допомогою гелікази без додаткового обертання ниток по відношенню один до одного повинно призводити до утворення на ділянках батьківської молекули перед рухається виделкою реплікації нових витків (вузлів), що створюють на них підвищений топологічний напругу. Така напруга усувається ще одним білком (ДНК-топоізомеразою), який, переміщаючись уздовж двухспіральной батьківської ДНК перед виделкою реплікації, викликає тимчасові розриви в одній з ланцюжків молекули, руйнуючи фосфодіефірні зв'язку і приєднуючись до розірваному кінця. Виниклий розрив забезпечує подальше обертання нитки подвійної спіралі, що, в свою чергу, призводить до розплітанню утворюються супервитки (вузлів). Оскільки розрив полі- нуклеотидной ланцюжка, викликаний топоізомеразою, має оборотний характер, то розірвані кінці швидко возз'єднуються відразу після руйнування комплексу цього білка з розірваним кінцем.
На другому етапі відбувається матричний синтез нових (дочірніх) Поліну клеотідних ланцюгів на основі принципу комплементарного відповідності нуклеотидів старої (матричної) і нової ланцюгів. Цей процес здійснюється шляхом з'єднання (полімеризації) нуклеотидів нового ланцюга за допомогою ферментів ДНК-полімерази декількох типів (ферменти Pol a, Pol р, Pol у, Pol 6, Pol е). Слід зазначити, що жодна з відомих сьогодні ДНК-полімерази не здатна почати синтез нового полинуклеотида шляхом простого з'єднання двох вільних нуклеотидів. Ініціація цього процесу вимагає наявності вільного -кінця будь-якої полинуклеотидной ланцюжка ДНК (або РНК), яка з'єднана з іншого (комплементарної) ланцюжком ДНК. Іншими словами, ДНК-полімераза здатна лише додавати нові нуклеотиди до вільного -кінцю наявного полинуклеотида і, отже, нарощувати цю структуру тільки в напрямку 5 * - *? 3 '.
З урахуванням зазначеної обставини стає зрозумілим асиметричний характер функціонування вилки реплікації (рис. 5.13, 5.14). Як видно з наведених схем, на одній з матричних ниток вилки (3 '- * 5') йде відносно швидкий і безперервний синтез дочірньої нитки (провідною, або лідируючої, ланцюжки) в напрямку 5 '-> 3', тоді як на іншій матриці (5 '-> 3') йде більш повільний і переривчастий синтез відстає ланцюжка короткими фрагментами (100-200 нуклеотидів), які отримали назву фрагментів Окадзакі, і також в напрямку 5 '-? 3'. Вважається, що синтез провідної і відстає ланцюжків здійснюють ДНК-полімерази різних типів. Вільний З'-кінець, необхідний для початку синтезу фрагмента Окадзакі, забезпечується короткою ниткою РНК (близько 10 нуклеотидів), що отримала назву РНК-праймера (РНК-затравки), яка синтезується за допомогою ферменту РНК-ПРАЙМАЗИ. РНК праймери можуть комплементарно спаровуватися відразу з декількома ділянками на матричної нитки ДНК, створюючи умови для одночасного синтезу декількох фрагментів Окадзакі за участю ДНК-полімерази Ш (рис. 5.14). Коли синтезований фрагмент Окадзакі досягає 5'-кінця чергового РНК-праймера, починає проявлятися 5'-екзонуклеаза активність ДНК-полімерази I, яка послідовно вищепляются нуклеотиди РНК в напрямку 5 * -? З *. При цьому видаляється РНК-праймер заміщається відповідним фрагментом ДНК.
Останній (третій) етап розглянутого процесу пов'язаний з дією ферменту ДНК-лігази, який з'єднує З'-ОН-кінець одного з фрагментів Окадзакі з 5'-Р04-кінцем сусіднього фрагмента з утворенням Фос фодіефірной зв'язку, відновлюючи таким чином первинну структуру відстає ланцюжка , що синтезується в функціонуючому РЕПЛІКОН. Подальша спирализация з'явився «напівконсервативним» ділянки ДНК (закручування спіралі) відбувається за участю ДНК-гірази (каталізує формування негативних супервитки в ДНК) і деяких інших білків.
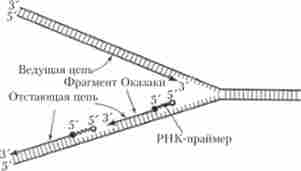
Мал. 5.14. Синтез провідною і відстає ланцюгів ДІК в області вилки реплікації
Полірепліконний принцип організації молекули ДНК різних еукаріот, в тому числі людини, забезпечує можливість послідовного копіювання генетичного матеріалу цих організмів без одночасного розкручування (деспіралізаціі) всієї величезної за розмірами і складно упакованої молекули, що значно скорочує час її реплікації.
Генетичний аналіз процесу реплікації ДНК у еукаріот в порівнянні з прокариотами виявив в основних рисах спільність синтезу ДНК у цих об'єктів. У клітині були виявлені і виділені в чистому вигляді ферменти, що володіють геліказной і топоізомеразной активностями, а також білки, специфічно зв'язуються з одноланцюжковий ділянками ДНК. Виявлено три види ДНК-полімеразної активності: ферменти Pol а (основна полімераза), Pol р (бере участь в репараційних процесах), Pol у (мітохондріальна полімераза).
Іншими словами, в той чи інший момент часу в одній групі репліконов молекули процес копіювання може бути вже завершений об'єднанням і спирализация відповідних ділянок, тоді як в іншій групі - тільки починатися розплітання двохниткових структур.