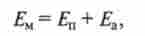
| Головна |
| «« | ЗМІСТ | »» |
|---|
Сприйняття і обробка інформації, що надходить із зовнішнього і внутрішнього середовища тваринного, починається в рецепторах. Розглянемо в загальних рисах будову і функціонування рецепторной частини аналізаторів, перш ніж перейти до викладу механізмів сприйняття рецепторами енергії дратівливих стимулів різних типів (різних модальностей) - механічних, світлових, хімічних, термічних.
Незважаючи на досить велику різноманітність рецепторів, що входять в різні аналізатори, їх можна розділити на дві групи, що підтверджують дані морфологічних досліджень, отримані на світловому та електронно-мікроскопічному рівнях. Рецептори першої групи представляють собою розгалужену термінальну частину нервового волокна (дендрита) першого чутливого, сенсорного, аферентного нейрона: всі ці назви позначають цю нервову клітину. Назва чутливий, або сенсорний, дано тому, що при порушенні рецептора в центральному відділі аналізатора тварини виникає відчуття. Однак в нормі робота аналізатора може відбуватися і без формування відчуття. Тому найбільш відповідна назва - аферентних нейрон, аферентних рецептор, т. Е. Що передає інформацію в ЦНС.
До першої групи належать аферентні рецептори шкірного аналізатора - механорецептори, реагують на механічне зміщення волосся, а також на дотик або натискання на безволосі ділянки шкіри; терморецептори, реагують на підвищення або зниження температури шкіри, - відповідно теплові або холодові; больові рецептори, реагують на сильні механічні і температурні впливи на шкіру і формують в центральному відділі відчуття болю. У цю ж групу входять і рецептори вісцерального аналізатора (внутрішніх органів): механо- і терморецептори, больові, а також реагують на зміну хімічного складу навколишнього середовища - хеморецептори.
Рецептори рухового аналізатора (пропріорецептори) - механорецептори м'язів і сухожиль також «утворені розгалуженнями термінальної частини першого аферентного нейрона. До першої групи належать і рецептори нюхового аналізатора, що сприймає молекули різних хімічних речовин - хеморецептори.
Друга група аферентних рецепторів представлена спеціальними клітинами, багато з яких на зверненої до дратівливої стимулу поверхні мають волоски. Перший аферентних нейрон пов'язаний з цими клітинами за допомогою хімічних синапсів. Такі рецептори складають початкова ланка слухового аналізатора і аналізатора положення тіла в просторі; вони реагують на механічні стимули, т. е. є механорецепторами. Видозмінені волоскові клітини являють собою рецептори зорового аналізатора, сприймають фотони світла - фоторецептори. І нарешті, рецепторні клітини смакового аналізатора, що реагують на хімічний склад їжі в ротовій порожнині у тварини, являють собою хеморецептори.
Необхідно відзначити, що термінальні частини рецепторів не прямим контактують з дратівливими факторами, а через навколишні їх структури, названі допоміжним апаратом. Допоміжний апарат має надзвичайно важливе значення для здійснення нормальної діяльності рецепторів: він може надавати модулюючий вплив на дратівливий стимул - посилювати чи послаблювати інтенсивність роздратування. Він же виконує бар'єрну функцію - обмежує вплив різних хімічних речовин і іонів з боку сусідніх клітинних структур на рецепторну область, зберігаючи навколо неї особливий склад середовища. Допоміжний апарат за своєю структурою значно варіює у різних рецепторів.
Порушення в нервовій системі може передаватися у вигляді двох типів електричних сигналів - градуальних і потенціалів дії, що генеруються за законом «все або нічого». Градуально потенціали, т. Е. Потенціали, що змінюють амплітуду в залежності від величини подразнення при найсприятливіших умовах проведення, поширюються на 2 ... 3 мм від місця свого виникнення (див. Гл. 2). Зв'язок між нервовими клітинами на більш тривалі відстані забезпечується тільки потенціалами дії. Беручи до уваги той факт, що аферентні рецепторні освіти у тварин знаходяться від ЦНС на відстані, рівному десяткам або навіть сотням міліметрів, енергія дратівної стимулу повинна трансформуватися в рецепторах в потенціали дії. Дійсно, при реєстрації електричної активності нервових волокон було виявлено, що в аферентних волокнах, що йдуть від різних рецепторів у центральній нервовій системі, крім потенціалів дії ніяких інших сигналів не генерується. З огляду на, що амплітуда потенціалів дії не змінюється, кодування інформації про дратівному стимулі в аналізаторах частотне. Це означає, що тривалі, безперервні впливу повинні перетворюватися в рецепторах в певним чином організовану послідовність потенціалів дії з різною частотою. Разом з тим вдосконалення методів реєстрації електричної активності, а також розробка методів експериментування з окремими нервовими клітинами і волокнами дозволили в середині XX в. наблизити місце реєстрації електричної активності безпосередньо до рецепторной області. Виявилося, що виникнення потенціалів дії в аферентних нервових волокнах передує градуальний електричний потенціал, що генерується в рецепторних закінченнях в результаті їх радраженія адекватними стимулами, що отримав назву рецепторного потенціалу. Нагадаємо, що мембранний потенціал нервових або м'язових клітин?м можна уявити як суму потенціалів:
де Е " - потенціал, що утворюється на мембрані за рахунок процесів пасивного транспорту іонів; Еа - потенціал, що утворюється на мембрані за рахунок активного транспорту іонів за умови, що іонний насос Електрогене.
З достатньою точністю?), Описується рівнянням Голдмана, з якого випливає, що мембранний потенціал клітини залежить як від різниці зовнішньої і внутрішніх концентрацій іонів, так і від їх проникності через мембрану. У свою чергу, мембранна проникність обумовлена числом відкритих іонних каналів. Для нервової і м'язової мембрани в спокої головним по- тенціалобразующім іоном є іон калію, для якого проникність мембрани найвища. При порушенні (генерації потенціалу дії) істотно збільшується в першу чергу проникність мембрани для іонів натрію, а потім для іонів калію. Обидва типи каналів є потенціалзалежні. Однак процес збільшення (активації) числа відкритих каналів в залежності від деполяризаціїмембрани для іонів натрію на відміну від калієвих каналів носить самоускоряющіхся (регенеративний) характер. З огляду на градуально рецепторного потенціалу, процес активації або інактивації іонних каналів, що обумовлюють його виникнення, не є регенеративним. Беручи до уваги різну модальність дратівливих стимулів, іонні канали, що знаходяться в області рецепторной мембрани, повинні мати в своїй структурі або ж поруч в мембрані ділянки (рецепторні білки), які можуть активуватися під впливом енергії адекватного для даного рецептора стимулу і відкривати або закривати канали.
На підставі вищевикладеного можна представити в загальному вигляді послідовність подій, що відбуваються в афферентной рецепторной клітці при дії на неї адекватного дратівної стимулу (рис. 15.1). Адекватний дратівливий стимул (/) через структури допоміжного апарату (2) впливає на мембрану рецептора. Енергія дратівної стимулу викликає зміни рецепторного білка (J), що знаходиться в клітинній
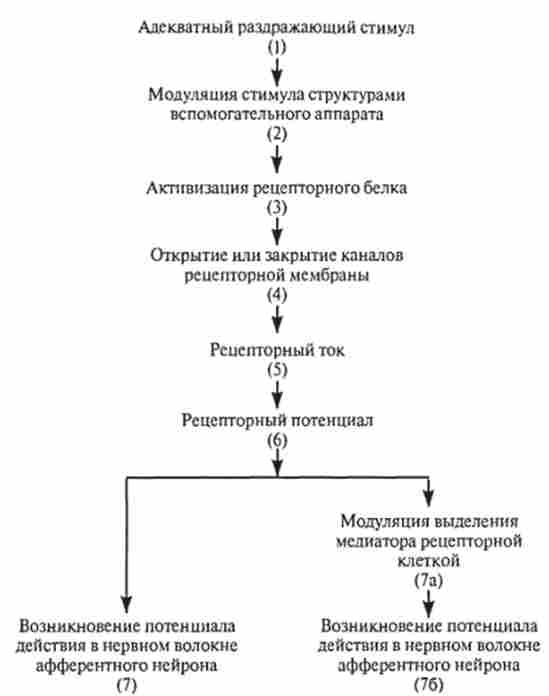
Мал. 15.1. Послідовність подій в афферентном рецепторі при сприйнятті дратівної стимулу
мембрані. Рецепторний білок або сам може бути частиною іонного каналу, або знаходиться поруч з каналом і здатний модулювати активність каналу опосередковано через ферментативні реакції. В результаті в обох випадках дія енергії дратівної стимулу на рецепторну молекулу приводить до відкриття або закриття іонних каналів (4), за якими проходить іонний струм (5). Рецепторний іонний струм змінює потенціал рецепторной мембрани. Ця зміна потенціалу, яка отримала назву рецепторного потенціалу (6), призводить до виникнення іонного струму, який викликає зрушення мембранного потенціалу в сусідніх ділянках нервового закінчення або нервової клітини. У першому випадку (7), якщо іонний струм буде деполярізуется, він може змінити мембранний потенціал до порогового рівня і викликати генерацію потенціалів дії, які будуть поширюватися по нервовому волокну в ЦНС. У другому випадку (7а) в результаті зміни мембранного потенціалу в синаптичної області зміниться рівень виділення медіатора. Якщо вихід медіатора збільшиться, в постсинаптичної області закінчення (76), утвореного аферентним волокном, відбудеться генерація потенціалу дії, який, як і в першому випадку, по нервовому волокну буде поширюватися в центральний відділ відповідного аналізатора. Слід зазначити, що через методичних труднощів рівень вивченості процесів трансформації енергії адекватного стимулу істотно відрізняється у різних рецепторів. Тому для ряду рецепторів деякі етапи представленої схеми ще мало вивчені. У наступних розділах ми більш детально ознайомимося з властивостями рецепторів різних модальностей.