| Головна |
| «« | ЗМІСТ | »» |
|---|
Все різноманіття рухів у тварин координується за допомогою двосторонніх зв'язків між ЦНС і м'язами тіла. Початкова ланка рухового аналізатора представлено сукупністю рецепторів, головним чином механорецепторов, які локалізуються в м'язах, сухожиллях, міжкісткових мембранах, фасціях, тканинах суглобів, окістя і т. Д. Тут можна виявити рецепторні освіти, зустрічаються і в інших частинах тіла тварини. Однак головними рецепторами рухового аналізатора є м'язові веретена і сухожильні рецептори Гольджі.
М'язові веретена належать до групи найбільш вивчених рецепторів. Тому слід більш детально розглянути їх характеристики з метою подальшого їх використання при вивченні властивостей рецепторів інших модальностей. Під світловим мікроскопом м'язове веретено є витягнуту структуру, розширену в середині за рахунок капсули і нагадує за формою веретено, що і зумовило його назву. М'язові веретена у всіх видів тварин мають приблизно однакову будову (рис. 15.2).
Допоміжні структури м'язових волокон представлені спеціалізованими поперечносмугастих м'язовими волокнами, названими інтрафузап'нимі (На відміну від звичайних, екстрафузальних волокон), а також капсулою рецептора, за-
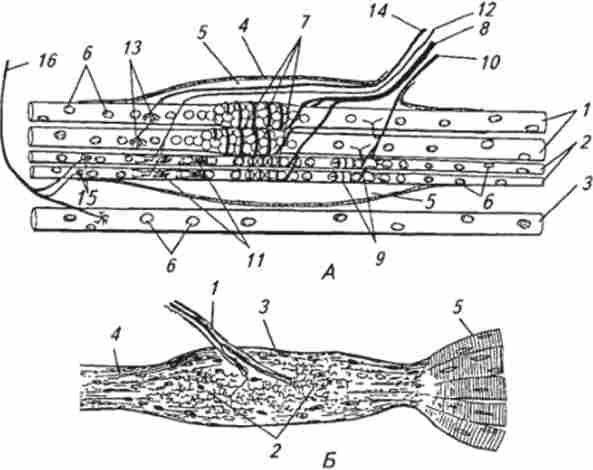
Мал. 15.2. Будова м'язового веретена (Л) і сухожильного рецептора Гольджі (Б):
А. 1 - інтрафузальних м'язове волокно з ядерної сумкою; 2 - інтра- фузальнос м'язове волокно з ядерної ланцюжком; 3 - екстрафузальнос м'язове волокно; 4-капсула; 5-внугрікапсулярное простір; 6-ядра м'язових волокон; 7-первинні аферентні закінчення; 8- афферентное нервове волокно першого типу; 9 - вторинні нервові закінчення; 10 афферентное нервове волокно другого типу; 11-у-кус тіковідние еферентні нервові закінчення; 12- Уг-рухове нервове волокно; 13 - у-пластінчатис еферентні нервові закінчення; 14- Yi-рухове нервове волокно; 75 - ^ -пластінчатие еферентні нервові закінчення; 16- По-рухове нервове волокно; Б. 1 - афферентное нервове волокно; 2 аферентні нервові закінчення; 3 капсула; 4-сухожилля; 5-м'язові волокна
конання рідиною. У полярних областях інтрафузальних волокон добре видно ядра, розсіяні по периферії волокна, як і в екстрафузальних. Ближче до центральної частини, де волокно покрито капсулою, з'являються додаткові ядра. Це і послужило підставою називати центральну зону волокна областю ядерної сумки, а самі інтрафузальних волокна називають волокнами з ядерної сумкою; діаметр цих волокон становить 20 ... 25 мкм.
У більш тонких волокон іншого типу (діаметр близько 10 мкм) ядра розташовуються в ряд, утворюючи ланцюжок. Такі волокна отримали назву інтрафузальних волокон з ядерної ланцюжком. Інтрафузальних волокна покриті сполучнотканинною капсулою. У центральній області веретена, де капсула відходить від інтрафузальних волокон, є перікапсулярное простір, заповнений рідиною. До м'язовому веретену підходить велика кількість нервових волокон, що розгалужуються в різних його частинах, але основна мережа розгалужень знаходиться в області, покритої капсулою. В м'язових веретенах було виявлено кілька типів аферентних і еферентних рухових закінчень. Останні відповідно до загальної схемою будови аналізаторів можна віднести до ланки зворотного зв'язку.
Аферентні закінчення представлені двома типами. Перший тип (первинні закінчення) утворюється розгалуженнями досить товстих міелінізірованних волокон, що досягають в діаметрі близько 20 мкм. Нервове волокно проникає під капсулу і, багаторазово розгалужуючись, поступово втрачає миелиновую оболонку. Первинні закінчення локалізуються в середині центральної ділянки інтрафузальних волокон і мають вигляд спіралей і кілець, що обвивають волокна. Другий тип (вторинні закінчення) утворюється більш тонкими міелінізі- рова волокнами (діаметр 4 ... 12 мкм) і розташовується ближче до периферії інтрафузальних волокна (див. Рис. 15.2). Діаметр обох типів закінчень в поперечному перерізі становить 0,5 ... 3 мкм. Характерна їх особливість - це велика кількість мітохондрій у внутрішньоклітинному просторі. М'язові веретена іннервують також і кілька типів рухових нервових волокон, які отримали назву фузімоторних (У-волокон). Фузімоторние синаптичні закінчення, утворені цими волокнами, підрозділяються на у-кустіковід- Цінні та у-пластинчасті. Перші локалізуються ближче до центру веретена, а у-пластинчасті - дистальніше. Крім рухових у-волокон м'язові веретена додатково іннервують рухові (3-волокна, які одночасно утворюють синаптичні закінчення на екстрафузальних м'язових волокнах. (3-волокна на інтрафузальних волокнах формують закінчення пластинчастого типу, розташовані в самих дистальних областях інтрафузальних волокон. У дослідженнях реакції інтрафузальних м'язових волокон на роздратування еферентних волокон було виявлено, що здатність до скорочення мають тільки дистальні ділянки, а центральна частина нез ритмами.
При реєстрації електричної активності в аферентних нервових волокнах, що йдуть від м'язових веретен, в стані спокою були виявлені нерегулярні низькочастотні потенціали дії, які отримали назву спонтанних. Беручи до уваги механізми генерації потенціалів дії в нервових клітинах і волокнах (див. Гл. 2), можна вважати, що мембрана рецепторной області від початку деполяризована і значення мембранного потенціалу знаходиться в області порога виникнення потенціалів дії. Таким чином, навіть найменше збільшення енергії дратівної стимулу (в даному випадку механічного) буде підвищувати частоту імпульсації рецептора і його чутливість дуже висока.
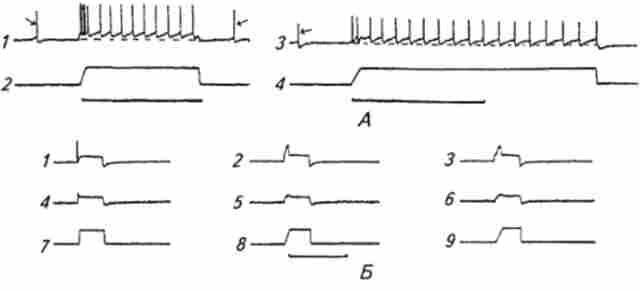
Мал. 15.3. Електричні відповіді м'язового веретена на ступенчатообразное механічне розтягнення:
А. / - електричні відповіді первинних аферентних нервових закінчень; 2 - відмітка ступенчатообразного механічного розтягування (калібрування часу: внизу горизонтальна лінія відповідає 0,5 с); 3 - електричні відповіді вторинних аферентних нервових закінчень; 4 - відмітка ступенчатообразного механічного розтягування (калібрування часу: внизу горизонтальна лінія-0,5 с); стрілками відзначені спонтанні потенціали дії. Б. Потенціали дії заблоковані введенням в розчин родотоксіна. Рецепторні потенціали первинних (/, 2, 3) і вторинних (4, 5, 6) аферентних нервових закінчень м'язового веретена у відповідь на розтягнення ступенчатообразнимі механічними стимулами з різною швидкістю наростання (7, 8, 9) переднього фронту механічного розтягування; калібрування часу: горизонтальна лінія
відповідає 0,5 с
Розтягування м'язового веретена призводить до виникнення електричної активності - рецепторного потенціалу і потенціалів дії (рис. 15.3). При легкому ступенчатообразном розтягуванні м'язового веретена в первинних і у вторинних аферентних закінченнях реєструється тривала серія потенціалів дії. Частота розрядів залежить від ступеня розтягнення м'яза. На малюнку видно, що потенціали дії знаходяться на своєрідному деполяризационного п'єдесталі, що представляє собою рецепторний потенціал, т. Е. Зміна мембранного потенціалу аферентних закінчень. Більш сильне розтягнення призводить до більш значної деполяризації і збільшення частоти аферентних потенціалів дії. Частота потенціалів дії м'язового веретена на тлі незмінної амплітуди розтягування змінюється з часом. Початок розтягування супроводжується підвищенням частоти імпульсів з наступним швидким спадом до певного рівня (динамічна частина відповіді). Далі частота продовжує вже повільніше знижуватися до значення, обумовленого величиною розтягування (статична частина відповіді). Процес зниження частоти потенціалів дії отримав назву адаптації. Таким чином, у відповіді м'язового веретена чітко виражені динамічна і статична фази, під час яких відповідно відбувається швидка і повільна адаптація. Подібна реакція на тривалий адекватний стимул характерна і для рецепторів інших модальностей. Даний тип рецепторів отримав назву медленноадаптірующіхся. Необхідно відзначити, що явище адаптації в аналізаторах має надзвичайно важливе значення для процесу сприйняття дратівної стимулу. Відповідно до закону специфічних енергій, зміни в частоті імпульсної активності аферентного нейрона показує переміну відчуття, що виникає в ЦНС. Робота рухового аналізатора у людини в нормальних умовах не супроводжується виникненням яких-небудь певних відчуттів. Непрямі дані вказують на те, що і у тварин руховий аналізатор функціонує подібним чином. Тому адаптація рецепторів м'язів не виявляється у відчутті. Однак в інших аналізаторах явище адаптації досить помітним чином впливає на сприйняття роздратування. Наприклад, в зоровому, нюховому, шкірному анализаторах відповідно інтенсивність відчуття яскравого світла, сильного запаху або низької температури з часом знижується.
Вивчення механізмів адаптації в рецепторах різних аналізаторів показало, що адаптація може відбуватися на різних етапах сприйняття роздратування (див. Рис. 15.1): від впливу стимулу на допоміжний апарат до генерації аферентних потенціалів дії. Причому у різних рецепторів внесок в адаптацію структур, що входять до їх складу, неоднаковий. Першою ланкою, яка визначає адаптацію у деяких рецепторів, служить допоміжний апарат. У м'язових веретен це інтрафузальних м'язові волокна. Як відомо (див. Гл. 2), м'язові волокна включають в себе еластичний і в'язкий елементи, по-різному реагують на розтягнення. Під дією розтягування спочатку відбувається подовження центральній частині з відповідною реакцією на це аферентних закінчень даної області, а потім дистальних ділянок веретена з роздратуванням контактують з ними аферентних закінчень. При цьому довжина центральної частини зменшується. Відповідним чином змінюються амплітуда рецепторного потенціалу і частота аферентних потенціалів дії в нервових волокнах. Наприклад, в первинних закінченнях, розташованих переважно в центрі веретена, за початковим підвищенням рецепторного потенціалу і частоти потенціалів дії слід швидке зниження (швидка адаптація). Надалі натяг буде послаблюватися повільніше і частота потенціалів дії почне знижуватися також повільніше (повільна адаптація). Паралельно з чисто механічним фактором в адаптації м'язового веретена беруть участь і іонні процеси, що лежать в основі генерації рецепторного потенціалу.
Порівняльні дослідження реакції первинних і вторинних аферентних закінчень м'язових веретен показали, що на стаціонарну частину розтягування вони реагують приблизно однаково
(Див. Рис. 15.3). Однак на зростання розтягування на початку дії стимулу відповіді їх відрізняються. Як видно на малюнку, у первинних закінчень динамічна частина відповіді більш виражена. Із зростанням швидкості розтягування збільшуються амплітуда динамічної частини рецепторного потенціалу і відповідно частота потенціалів дії, що припадають на цей період. У вторинних закінчень динамічна частина відповіді виражена в меншій мірі. Таким чином, вторинні аферентні закінчення головним чином кодують (детектируют) амплітуду розтягування - детектори амплітуди, а первинні закінчення - амплітуду і швидкість наростання розтягування - детектори швидкості амплітуди.
При введенні в розчин, що омиває м'язове веретено, тет- родотоксіна - специфічного блокатора потенціалзавісімих натрієвих каналів (див. Гл. 2) - потенціали дії придушуються. Однак рецепторний потенціал, який має ступенчатообразную форму з характерним деполяризационного викидом, на початку свого виникнення (динамічна частина відповіді) зберігається і не змінюється з часом. Це вказує на різні механізми генерування рецепторного потенціалу і потенціалів дії, що виникають згідно із законом «все або нічого». На цьому ми зупинимося докладніше далі. Посилення інтенсивності адекватного стимулу (розтягування) супроводжується підвищенням як амплітуди рецепторного потенціалу, так і частоти аферентних потенціалів дії. Діапазон інтенсивностей стимулу, в якому рецептор здатний кодувати амплітуду стимулу, генеруючи зі збільшенням енергії стимулу все більш інтенсивні сигнали, називається динамічним діапазоном.
В м'язових веретенах амплітуда рецепторного потенціалу приблизно пропорційна логарифму інтенсивності стимулу (рис. 15.4.) У той же час середня частота аферентних імпульсів пов'язана більш-менш лінійною залежністю з амплітудою рецепторного потенціалу аж до того значення, ко-
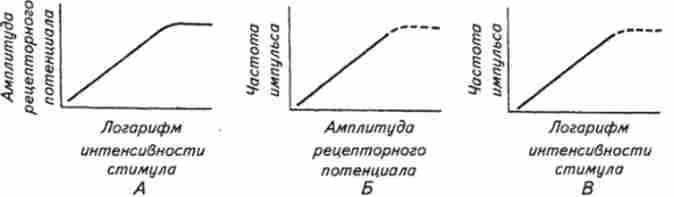
Мал. 15.4. Залежність між амплітудою рецепторного потенціалу і логарифмом величини розтягування (Л), частотою імпульсів і амплітудою рецепторного потенціалу (Б), Частотою імпульсів і логарифмом величини розтягування (В) в м'язових веретенах
да явище рефрактерності потенціалів дії (див. гл. 2) почне обмежувати інтервал між потенціалами дії і, отже, частоту імпульсації. Виходячи з цих двох залежностей, частота потенціалів дії є функцією логарифма інтенсивностей стимулу в медленноадаптірующіхся рецепторах. З огляду на той факт, що ступінь розтягнення м'яза може змінюватися в досить широких межах, стає зрозумілим, наскільки важлива для м'язових веретен логарифмічна залежність між амплітудою стимулу і частотою аферентних потенціалів дії. Ця залежність також має надзвичайно велике значення для сприйняття роздратування у рецепторів інших модальностей. Так, наприклад, сонячне світло приблизно в 1019 раз яскравіше місячного, а слухова система людини і тварин здатна сприймати без значного спотворення звуки, енергія яких може різнитися в 1012 раз. Тут треба зазначити, що здатність аналізаторів функціонувати в великих діапазонах інтенсивності зовнішніх стимулів визначається не тільки великим діапазоном первинного (рецепторного) перетворення стимулів, але і наявністю в центральних структурах аналізатора механізмів, додатково розширюють динамічний діапазон. Логарифмічна залежність «роздратування - відповідь» (див. Рис. 15.4) «стискає» область високих інтенсивностей, тим самим значно розширюючи діапазон, в якому можливо розрізнення стимулів за амплітудою. Внаслідок логарифмічною залежності між величиною рецепторного потенціалу і силою роздратування дане відносне посилення стимулу призводить до однакового приросту рецепторного потенціалу в дуже широкому діапазоні інтенсивностей. Наприклад, подвоєння сили слабкого дратівної стимулу призведе до такого ж приросту амплітуди рецепторного потенціалу, як і при подвоєнні сили стимулу в області високих інтенсивностей. Якщо позначити інтенсивність стимулу /, а приріст стимулу Л /, то відношення приросту стимулу до його амплітуді буде величиною постійною, т. Е.
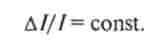
Яка ж природа рецепторного потенціалу м'язових веретен? Експерименти на м'язових веретенах з варіацією іонного складу в зовнішньому розчині показали, що рецепторний потенціал генерується в основному за рахунок іонів натрію. Однак в розчині з тетродотоксином рецепторний потенціал не пригнічується (див. Рис. 15.3, Б). Це свідчить про те, що натрієві канали в рецепторной мембрані мають іншу структуру, ніж потенціалзалежні натрієві канали нервових волокон. Мабуть, їх ворітної механізм відрізняється від такого в потенціалзавісімих натрієвих каналах. Активація його відбувається через конформаційні (просторові) зміни, викликані механічним впливом. Такі канали отримали назву механоактівіруемих. На жаль, методичні труднощі не дозволяють поки вивчати поодинокі канали рецепторной мембрани м'язових веретен. Однак вдалося за допомогою методу локальної фіксації (див. Гл. 2) зареєструвати відповіді механоактівіруемих іонних каналів в мембрані механорецепторних дендрита м'язового рецептора розтягування річкового рака. Виявилося, що іонний струм через одиночний механоактівіруемий канал, як і в випадку з потенціалзалежні і ацетілхолінактіві- руемой каналами (див. Гл. 2), має ступенчатообразную форму різної тривалості. Зі збільшенням розтягування мембрани ймовірність відкритого стану каналу підвищувалася, причому залежність між інтенсивністю дратівної стимулу і ймовірністю відкриття каналу була логарифмічною.